- Creation
-
Creator (Definite): Sandra Elizabeth BlackDate: 1981
- Current Holder(s)
-

- No links match your filters. Clear Filters
-



-
 CitesF.X. Dercum, 'The Functions of the Neuron', Journal of Nervous and Mental Diseases 23 (8) (1896), pp. 513-523.
CitesF.X. Dercum, 'The Functions of the Neuron', Journal of Nervous and Mental Diseases 23 (8) (1896), pp. 513-523.
Description:'The American neurologist, Francis Xavier Dercum (1856-1931) in his presidential address to the American Neurological Association in 1896, [note: 'F.X. Dercum, "The functions of the neuron," J. Nerv. Ment. Dis., 1896, 21: 518-23.'] heralded the amoeboid hypothesis as the new basis for a "rational and biological psychology," and showed how it could elucidate many clinical features of hysteria and hypnosis.' (45)
-
 Cited by
Cited by Henri Bergson's Physiological Psychology: Vitalism and Organicism at the Start of the Twentieth Century
Henri Bergson's Physiological Psychology: Vitalism and Organicism at the Start of the Twentieth Century
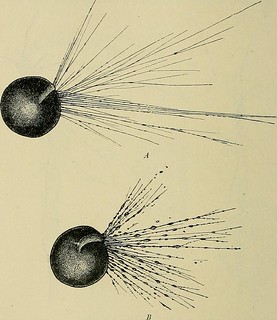
Description:'Exactly how independently-acting cellular individuals could combine to form more complex psychological wholes [was uncertain around 1900]. Different tendencies existed within the conceptual framework of cellular individuality. One contention that gained considerable traction during the final years of the nineteenth century was the so-called 'amoeboid' theory of cellular interaction. Just as individual cells could be identified as manifesting vital capacities such as autonomous extension and contraction, each nerve-cell or 'neuron' was, in the conception of histologists such as Mathias-Marie Duval, an independently-acting contributor to the neurological whole. Ramón y Cajal suggested in a study of 1890 that nerve fibres could be seen to grow outwards from their cellular origins. Though Cajal would later renounce this view, he and like-minded theorists believed that this constituted evidence that nerve cells behaved just like amoeba in that they expanded, contracted, and moved their extremities from place to place.'
'Adherents of cellularly-distinct conceptions of neuronal connection portrayed them as confirming associationist contentions regarding the nature of psychology. For example, US physician Francis Xavier Dercum argued that the 'amoeboid' expansion and retraction of individual cells underlay variously: hysteria; hypnotic and dream states; sleep; and trains of thought themselves. These latter, Dercum contended, appeared 'to follow purely mechanical lines' of association and disassociation between the sense impressions that they carried. In 1898, Boris Sidis and Ira van Gieson separately elaborated this thesis to account for states of sanity and insanity: in Gieson's words, 'unsoundness of mind' was caused by the 'dissociation' of the 'higher and last evolved parts of the brain, in the presence of pathogenic stimuli.' For amoeboid theorists, the apparent mechanical capacity of nerve cells to alter their proximity to one another presented physical confirmation of the existence of psychological associations.'
-
Cited byR.M. Brain, 'Materialising the Medium: Ectoplasm and the Quest for Supra-Normal Biology in Fin-de-Siècle Science and Art', in A. Enns and S. Trower (eds.), Vibratory Modernism (Basingstoke: Palgrave Macmillan, 2013), pp. 115-144.
Description:'In the 1890s the nerve junction was still understood through the amoeboid theory of neuronal mobility, which held that neurons made functional contacts through pseudopodal movements of the protoplasm of nerve cells. [note: 'Sandra E. Black, 'Pseudopods and Synapses: the Amoeboid Theories of Neuronal Mobility and the Early Formation of the Synapse Concept. 1894-1900', Bulletin of the History of Medicine 55 (1) (1981): 34-58.']' (12)
-
Cited byT. Quick, 'Disciplining Physiological Psychology: Cinematographs as Epistemic Devices, 1897-1922', Science in Context 30 (4), pp. 423-474.
Description:'Ramón y Cajal, whose work Sherrington both promoted and came to rely upon, suggested in a study of 1890 that nerve fibres could be seen to grow outwards from their cellular origins. He and like-minded theorists believed at this time that such observations constituted evidence that nerve cells behaved just like amoeba, in that they expanded, contracted, and moved their extremities from place to place (Black 1981, on 35-36 and 38-39).'
'Adherents of cellularly-distinct conceptions of neuronal connection portrayed such studies as confirmatory of associationist contentions regarding the nature of psychology (Black 1981, 44-45). For example, US physician Francis Xavier Dercum argued that the 'amoeboid' expansion and retraction of individual cells underlay variously: hysteria; hypnotic and dream states; sleep; and trains of thought themselves. These latter, Dercum contended, appeared 'to follow purely mechanical lines' of association and disassociation between the sense impressions that they carried (Dercum 1896, esp. 520-523. Quote on 522. Original emphasis). In 1898, Boris Sidis and Ira van Gieson elaborated this thesis to account for states of sanity and insanity: in Gieson's words, 'unsoundness of mind' was caused by the 'dissociation' of the 'higher and last evolved parts of the brain, in the presence of pathogenic stimuli' (van Gieson 1899, 87; Sidis 1919, 208-215). For amoeboid theorists, the apparent mechanical capacity of nerve cells to alter their proximity to one another presented physical confirmation of the existence of psychological associations.'
'In his Textbook of Physiology of 1900, Edward A. Schäfer had suggested that impulses were 'momentarily arrested at these places of contact of nerve cells with one another', which he referred to as 'synapses' (Schäfer 1900, 608). Sherrington, characterizing such arrests as manifestations of a 'neurone threshold', identified it unequivocally with a non-contiguous, synaptic transmission:
at each synapse a small quantity of energy... acts as a releasing force to a fresh store of energy not along a homogeneous train of conducting material, as in a nerve fibre... but across a barrier which whether lower or higher is always to some extent a barrier (Sherrington [1906] 1947, 157). [note: 'On Sherrington's earlier 'reticulist' conception of synapses see Black 1981, 55-56.']'
Relevant passages from Black:
'amoeboid theories were put forward by French and Belgian histologists between 1894 and 1900, and elicited responses from the leading histologists of the day. The ensuing debate was also pursued outside histological circles, though it did not engage the psychologists, who may have found the theories too simplistic. [note: 'See, for example, L. Azoulet, "Psychologie histologique et texture du système nerveux," L'Année Psych., 1895, 2: 255-56, 292-94.'] The general framework within which the amoebists attempted to provide histological explanations of psychological phenomena was association psychology, the theory of mental function that predomintated in the second half of the nineteenth century. ['for a discussion of the association theories being promulgated in this period by such influential thinkers as Alexander Bain, Herbert Spencer, Wilhelm Wundt and others, see E.G. Boring, A History of Experimental Psychology, 2nd ed. (New York: Appleton-Century-Crofts, 1950).'] With the advent of neuron theory, there was a tendency among some histologists to assume that the neurological basis of the temporal and spatial association of ideas was quite literally the "association " of neurons through interneuronal conduction. [note: 'See, for example, S. Ramon y Cajal, "Einige Hypothesen ueber der anatomischen Mechanismus der Ideenbicklung der Association und der Aufmerksamkeit," Arch. f. Anat. Physiol., 1895, suppl.: 367-78.'] Alterations in interneuronal conduction, for which the amoebists proposed a mechanism, were thought to underlie alterations both in the state and in the content of consciousness. The particular problem in clinical psychology to which amoebists addressed themselves were hysteria and hypnosis, the two subjects being eagerly studied by medical psychologists in the 1880s and 1890s. [note: 'For accounts of clinical psychology in this period, see G. Zilboorg, A History of Medical Psychology (New York: W.W. Norton, 1941) and E.H. Ackernecht, A Short History of Psychiatry, Tr. by S, Wolff, 2nd ed. (New York: Hafner, 1968)'] But the main feature of the Amoeboid hypothesis as an explanation of psychologicla phenomena was that it was pre-eminently biological, rooted in the two main biological generalizations of the nineteenth century - cell theory (1839), and evolutionary biology (1859). [note: 'For a discussion of the development of cell theory, which was first enunciated by Theodor Schwann in 1839,see A. Hughes, A History of Cytology (New York: Abelard-Schuman, 1959). The impact of Darwin's evolutionary thory is discussed in A.C. Seward, ed., Darwin and Modern Science (Cambridge: Cambridge University Press, 1909), and S.A. Benett, A Century of Darwin (London: Mercury Books, 1962).]
The complexity of the nervous system and the need to develop suitable histological techniques, meant that it took fifty years for cell theory to be verified for nervous tissue. Several essential steps in the development of the neuron concept can be identified. [note: 'For details see E.G.T. Liddell, The Discovery of Reflexes (Oxford: Oxford University Press, 1960), pp. 1-30; A. Andreoli, Zur geschichlichen Entwicklung der Neurontheorie (Basle: B. Schwabe, 1961); H. van der Loos, "The history of the Neuron," in H. Hyden, ed. The Neuron (Amsterdam: Elsevier, 1961), pp. 5-47; E. Clarke and C.D. O'Malley, The Human Body and Spinal Cord (Berkeley and Los Angeles,: University of California Press, 1968), pp. 27-138.'] Nerve cell bodies nad nerve cell fibres in the central nervous system were first described in the 1830s, but were assumed to be separate elements. Evidence that fibres and cell bodies were connected was provided in the 1840s, though this idea was not widely accepted until the 1860s. The two kinds of nerve branches - axons and dentrites - were first differentiated by the german histologist, Otto Deiters (1834-1863), in a monograph published posthumously in 1865. [note: 'O. Deiters, Untersuchungen ueber Gehirn und Rüchenmark des Menschen und des Saugethiere (Braunschweig: F. Vieweg, 1865.)'] For many years it was believed that the branches formed a continuous network in the central grey matter, but in 1883 the Italian histologist Camillo Golgi (1843-1926), usng his silver impregnation technique, demonstrated that dentrites end freely and and not in continuity with other nerve endings. [note: 'C. Golgi, "Recherches sur l'histologie des centres nervaux," Arch. ital. biol., 1883, 3: 285-317; 1884, 4: 92-123. Golgi's important technique, discovered in 1873, only became widely known after Golgi, who remained an opponent of neuron theory, shared the Nobel prize with Cajal in 1906.'] he held that nerve conduction occurred only through axons and their collaterals, which formed a continuous network in the grey matter, and tuhat the dentrites had a non-conductive, nutritional function. In 1887 the German neuroembryologist, Wilhelm His (1831-1904), and the Swiss neuropsychiatrist August Forel (1848-1931), suggested that axon terminals also end freely. [note: 'W. His, "Zur Geschichte des menschlichen Rüchenmarks und den Nervewurzeln," Abb. k. sach. Ges. Wiss., Math-Phys. Classe, 1887, 13;: 477-514 (dated 1886); A. Forel, Einige hirnanatomische Betrachtungen und Ergebnisse," Arch. f. Psychiat. Nervenkrankh, 1887, 18: 162-98.'] but the first to give direct histological evidence for this was the Spanish histologist, Ramón y Cajal (1852-1934). [note: 'S. Ramón y Cajal, Recollections of my Life, Tr. by E.H. Craigie and J. Cano (Cambridge, Mass: MIT Press, 1937), pp. 328-32. Cajal used a modified Golgi method to examine the cerebellum in young animals. See S. Ramón y Cajal, Les Nouvelles Idées sur la structure du système nervaux, Tr. by L. Azoulet (Paris, C. Reinwald, 1895) for a summary of the early researches of Cajal that were fundamental to the establishment of neuron theory. The impressive contributions to neurohistology of this legendary Spanish investigator are well described in Cajal's Recollections, and in D.F. Cannon, Explorer of the Human Brain: The Life of Santiago Ramón y Cajal (1852-1934) (New york: H. Schuman, 1949).'] His work, published in 1888, was soon verified by others, [note: 'For example, Albert von Kölliker, Gustav Retzius, and Arthur van Gehuchten. See L.F. Barker, The Nervous System and its Constituent Neurones (New York: D. Appleton, 1899), pp. 20-30, and Cajal, Recollections, pp. 352-62.'] and the nerve cell, with its free ending terminations, was at last recognized as a discrete anatomical unit, forst called the "neuron" in 1891 in an influential review by the German histologist, Heinrich Waldeyer, (1837-1921). [note: 'H.W.G. Waldeyer, Ueber einege neuere Forschungen im Gebiete der Anatomie des Centralnervensystems," Deutsche Med. Wschr, 1891, 17: 1213-18, 1244-46, 1267-69, 1287-89, 1331-32, 1352-56.']' (35-36)
'The extension of this concept [of chemiotaxis] to the explanation of nervous activity was first applied to embryonic nerve cells. In 1886, His had published new evicdence for the "outgrowth theory" of axonal development, according to which each axon "originates as a process from a single cell." [note: 'His, Geschichte des Rückenmarks, p. 514. See also W. His, "Ueber die embryonale Entwickelung der Nervenbahen," Anat. Anz, 1888, 3: 499-506.'] First advanced in 1857, this theory had been less popular than its rival, the "multicellular theory," which postulated that an axon was formed either within, or as an extracellular product of, a pre-existing chain of cells that extended from the axon's cell of origin to its destination. [note: 'For an account of these theories, see S.M. Billings, "Concepts of Nerve fibre Development, 1839-1930," J. Hist. Biol., 1971, 4: 275-305.'] His was unable, in his preparations, to see the actual tip of a nerve fibre as it grew out from its cell of origin, and this remained a drawback to "outgrowth theory" until 1890 when Cajal was able to demonstrate the growing tip of a nerve fibre in chick embryo using the Golgi silver impregnation method. [note: 'S. Ramón y Cajal, "A quelle époque appraissant les expansiones des cellules nerveuses de la moelle épinière du poulet?" Anat. Anz. 1890, 5: 609-13, 631-39.'] Cajal's visualization of this "growth cone" gave great impetus to outgrowth theory, [note: 'Lenhossék confirmed Cajal's discovery in 1892 and described the growth cone as a "mass of protoplasm, which by a kind of 'amoeboid movement' works its was though the tissue." M. von Lenhossék, Der feinere Bau der Nervensystems in Lichter neuester Forschungen, 2nd ed. (Berlin, H. Kornfeld, 1895), p. 94.'] but another problem was its difficulty in explaining how axons were able to reach their destination, often over long distances, from their cells of origin. The multicellular theory, with its concept of a pre-existing chain of cells to guide axonal growth, made this devlopmental feat more comprehensible. [note: 'It was not until 1907, when R.G. Harrison observed axons growing from neuroblasts in culture, that the outgrowth theory finally prevailed. See R.G. Harrison, "Observations on the living developing nerve fibre," Anat. Rec., 1907, 1: 116-18, and Billings, "Concepts of nerve fibre," pp. 295-302.'] Cajal's solution was to propose that the directed growth of nerve fibres represented a chemiotacticphenomenon. Citing the work of Metchnikoff and others on chemiotaxis in motile cells, he hypothesized in 1892 that neuroblasts also possess chemiotactic sensibility:
If one admits a chemiotactic sensibility in neuroblasts, one must suppose that these elements are endowed with amoeboid movement, and that they are excited by the subltances secreted by certain nerve, epithelial or mesodermal cells. The neuroblast elongations would be oriented in the direction of chemical currents and go to meet the secreting cells. [note: 'S. Ramón y Cajal, "La rétine des vertébréts," La Cellule, 1893, 9: 237 (dated 1892).']
Cajal employed the concept of chemiotaxis to explain more than just the outgrowth of nerve processes. It was already known that at certain embyonic stages neuroblasts actually move from one location in the nervous system to another. Cajal suggested that chemiotaxis lay at the root of these movements as well. This use of the concept of chemiotaxis to explain the growth and migration of embryonic nerve cells lent further credibility to the analogy between amoebae and adult neurons that underpinned the amoeboid theories.' (38-39)
'Cajal records that the Lépine-Duval hypothesis of neuronal mobility was initially received very sympathetically. [note: 'S. Ramón y Cajal, Histologie du système nerveux de l'homme et des vertébrés, Tr. by L. Azoulay, 2 vols. (Paris, Maloine, 1909-11), 2: 883. Interest in the new hypothesis brought to light an amoeboid theory proposed in 1890 (and subsequently ignored) by H. Rabi-Rückhard (1893-1905 [sic]) in the context of the old network theory. See H. Rabi-Rückhard, "Sind die Ganglienzellen amöboid? Eine hypothese sur Mechanik psychischer Vorgange," Neurol. Zbl., 1890, 7: 199-200 and A. von Kölliker, "Kritik der Hypothesen von Rabi-Rückhard und Duval ueber amoeboide Bewegungen der Neurodendren," Sitzungsb. Wurzburger Psychik-Medic. Geselsch., 1895: 38-42. Rabi-Rückhard suggested that the dendrites of the cortical pyramid cells formed a complex dynamic network which, during cerebral processes such as thinking, was continally breaking up and reuniting so as to produce a constant change in connections. The British histologist Alexander Hill, also in the context of network theory, raised the possibility that the "protoplasmic processes of nerve cells do not form a permanent network but have a certain mobility, forming valuable connections along the lines traversed by nerve impulses," in a review, "Current nerve anatomy and physiology," Brain, 1891: 568.'] The most enthusiastic responses came from outside histological circles. In a review of neuron theoryin 1896, [note: 'V.M. Bekhterev, "Die Lehre von den Neuronen und die Entladungstheorie," Neurol. Zbl., 1896, 15: 106-7. Bekhterev, who made substantial contributions to neurology and neuroanatomy, is best known for his later work on conditioned reflexes and social behaviour (which he called "Psychoreflexology") the Russian neuropsychiatrist, Vladimir Bekhterev (1857-1927), found the mobility hypothesis plausible and in harmony with his own speculations, but he awaited empirical support. [note: 'Another Russian psychiatrist, Serge Soulkhanhoff, combined features of the amoeboid hypothesis with Cajal's growth theory of learning to explain the neurological basis of mania, retarded depression and the organic brain syndromes. See S. Soulkhanoff, "Les théorie des neurons en rapport avec l'explication de quelques ètats psychiques normaux et pathologiques," Arch. de Neurol., 1897, 3: 337-47; 4: 15-28. The Italian psychiatrist, Ernest Lugaro, also utilized the amoeboid hypothesis and Tanzi's growth theory in his speculations on the neurological basis of various psychiatric syndromes. See E. Lugaro, "I Fatti i le induzione nell'odierna istologa del sistema nervoso," Ric. sperim, freniatria medic. leg., 1893, 19: 419-72.'] The American neurologist, Francis Xavier Dercum, (1856-1931), in his presidential address to the American Neurological Association in 1896, [note: 'F.X. Dercum, "The functions of the neuron," J. Nerv. Ment. Dis., 1896, 21: 518-23.'] heralded the amoeboid hypothesis as the new basis for a "rational and biological psychology," and showed how it could elucidate many clinical features of hysteria and hypnosis. The American neuropathologist, Ira van Gieson (1866-1913), was no less eager in a review published in 1898. As if the analogy to amoebae was not graphic enough, he spoke of the neuron as a "tiny octopus" with "the power of movement over its shorter arms or tentacles." [note: 'I van Gieson, "The correlation of sciences in research," J. Ment. Sci. 1898, 44: 768. Van Gieson discovered a special collagen staining technique in the 1890s which is still in use today.'] He applied the amoeboid hypothesis to the explanation of epilepsy, suggesting that a seizure resulted when some stimulus caused a sudden retraction of neuron groups in the association cortex and released the lower sensory and motor centres from inhibitory control.' (44-45)
-
-
-
-

-

