- External URL
- Creation
-
Creators (Definite): Marion Greenwood Bidder; Edith Rebecca SaundersDate: 1894
- Current Holder(s)
-

- No links match your filters. Clear Filters
-



-
-
 CitesÉ. Metchnikoff, 'Recherches sur la digestion intracellulaire', Annales de l'Institut Pasteur 3 (1889), pp. 25-29.
CitesÉ. Metchnikoff, 'Recherches sur la digestion intracellulaire', Annales de l'Institut Pasteur 3 (1889), pp. 25-29.
Description:'The possibility that the substance of animal cells may give rise within itself to accumulations of acid fluid, even when these cells are not highly differentiated, has been considered indeed by many earlier writers, and their statements have gained a degree of familiarity by repetition. But if we except a paper by M. Metchnikoff [note: 'E. Metchnikoff. Annales de l'Inst. Past. 1889, p. 25.'], which contains a discussion of the reaction of plasmodia to ingested litmus, we find that the records which bear on this question are often incidental and unsupported by the details of experiment.' (441)
'In point of time... , this work [of le Dantec] was preceded by the paper in which Metchnikoff describes the ingestion of blue grains of powdered litmus by different species of plasmodia and subsequent change of the blue colour to red, pointing out that, as the substance of the plasmodia is clearly alkaline, this change indicates the secretion of an acid fluid. He records a comparable result with certain of the larval phagocytes in the tail of Triton toeniatus, and, noting that in the same cell a reddened grain of litmus may coexist with a vacuole in which blue grains lie, he draws the inference that the acid secretion may be localized even in the relatively small mass of a single wandering cell.' (442-443)
'while Metchnikoff and le Dantec are alike in emphasizing the intracellular secretion of acid fluid by relatively undifferentiated protoplasm, neither of them deals at any length with the part played by this acid in the solution of nutritive material as it is carried out within the cell; the point which we, on the other hand, were most anxious to examine is the transformation undergone by acid indicators as associated with successive phases of the digestive process.' (444)
-
CitesH.A. de Bary, Vergleichende Morphologie und Biologie der Pilze, Mycetozoen, und Bacterien (Leipzig, 1884).
Description:'The possibility that the substance of animal cells may give rise within itself to accumulations of acid fluid, even when these cells are not highly differentiated, has been considered indeed by many earlier writers, and their statements have gained a degree of familiarity by repetition. But if we except a paper by M. Metchnikoff [note: 'E. Metchnikoff. Annales de l'Inst. Past. 1889, p. 25.'], which contains a discussion of the reaction of plasmodia to ingested litmus, we find that the records which bear on this question are often incidental and unsupported by the details of experiment. Among these the following statement made by De Bary [note: 'E. de Bary. [sic] Pilze, Mycet. und Bact. p. 487.'], and now almost classical, must be placed, - "As to the changes which ingested bodies may undergo in the interior of a plasmodium, this one observation is established, that, in the case of Didymium serpula, carmine is dissolved." No detailed account of the observation is given nor is there any attempt to harmonize it with the statement which succeeds immediately, that carmine is ingested rarely and never dissolved by the nearly allied Didymium libertianum. It seems clear that De Bary is concerned rather with the fact that solid bodies are ingested by the Mycetozoa than with the intracellular surroundings of ingesta.' (441)
'Since the days when De Bary and Engelmann wrote, observations have been made by one of us on the digestive process in Amoeba and Actinosphaerium. These observations sought to establish the constant association of a fluid medium with the manifestation of proteolytic activity in Protozoa, and secondarily, the reaction of this medium.' (442)
'Carmine grains ingested alone are unchanged by Carchesiurn or by the plasmodia we have used; ingested with nutritious matter they are equally unchanged, but they adhere to the lessening remnant of food throughout digestion. When ingesta stained with carmine as a preliminary to enclosure are watched, the digestive vacuole is seen to be colourless at first but soon afterwards is clearly red. This colour which belongs wholly to the solution of stained matter seems to us comparable to the red colour set free in peptic digestion of carmine-stained fibrin [note: 'Cp. P. Grützner, Neue unter. ü. d. Bild. u. Auss. d. Peps. (Breslau, 1875); and J. N. Langley, This Journal, Vol. III. 1880.']; it is in fact distinctly different from the "lake" colour of carmine in alkaline solution. We have examined these relations carefully and repeatedly because of the use made of carmine by De Bary in the experiments quoted at the beginning of this paper. It will be remembered that he speaks of the solution of carmine within the plasmodium of one mycetozoan, while a nearly allied form ingested little and dissolved none. Many succeeding writers have quoted De Bary's statement; Pfeffer [note: 'Op. cit.'] discusses it in the papers to which we have referred above, and we gather that he is inclined to explain the result by postulating the possibility of locally variable response to like stimuli from any area of a plasmodium. By such an hypothesis solution of carinine and inaction on carmine are demonstrable, conceivably, on neighbouring ingesta. No instance of such extreme local dissimilarity is adduced, but stating that Chondriodermoa difforme [note: 'The Didymium Libertianum of De Bary'] ingests carmine abundantly (in this form De Bary found that there was practically no ingestion) Pfeffer notes that in most cases the grains remained unaltered, while exceptionally, a vacuole, coloured red by solution, is formed. We think that the possibilities of variable reaction are somewhat overestimated by Pfeffer; for us, as formulated by him, they transcend experience. And bearing in mind how actually meagre is this classical statement, how partial the support which it has gained in later years [note: 'Cp. A. Lister. Journ. Linn. Soc. Vol. L. The ingestion of carmine grains by swarm cells of Mycetozoa is described without subsequent digestion (p. 439.)'], and the entire absence of solution of carmine grains which is noticeable constantly in our experiments, we think it a more probable hypothesis that solution (when it occurred) was due rather to some accidental constituent of a vacuole ingested from without than to secretion poured from the surrounding cell substance.' (457-458)
-
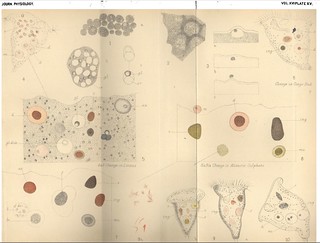 CitesPlate XV, Journal of Physiology 16 (5-6) (1894). Figs. 1-10 from M. Greenwood and E.R. Saunders, 'On the Rôle of Acid in Protozoan Digestion'.
CitesPlate XV, Journal of Physiology 16 (5-6) (1894). Figs. 1-10 from M. Greenwood and E.R. Saunders, 'On the Rôle of Acid in Protozoan Digestion'.
Description:Explanation of Plate XV (figs. 1-10):
'In all the figures
ing = a solid particle soon after enclosuire.
s = a solid particle "stored."
d = a solid particle during some period of actual digestion.
ex = a solid particle or a mucilaginous vacuole when digestion is practically complete.
m = initermittence of digestion with concentration of mucilaginous matter round solid. (See text p. 449.)
Fig. 1. a = Sclerotium cells of Mycetozoa. Oc. 2. Obj. D. Zeiss.
b = Endosperm cell of Ricinus communis, cell wall removed to show crystalloids and globoids in situ.
c = isolated aleurone grains of Ricinus communis, showing globoid intact, and dissolved. Oc. 2. Obj. F.
Fig. 2. Lime-holding plasmodium. Oc. 2. Obj. F.
Fig. 3. To show extrusion of undigested matter from vacuole. Oc. 2. Obj. F.
a = vacuole and food-mass present.
b = food-mass present after discharge of vacuole.
c = food-mass extruded.
Fig. 4. Ingestion of litmus and white of egg by Carchesium. Red colour is distinct in storage, and changes to blue during digestion.
[note: 'The red colour acquired by ingested litmus is really more yellow in tone than the shade of red used in fig. 5.'] Fig. 5. Ingestion of litmus in plasmodiun. Aleurone grains stained. Storage = red. Beginning of digestion = red. Advanced digestion = violet passing to blue.
Fig. 6. Ingestion of Congo red and white of egg by Carchesium; blue colour (transient) in storage stage.
Fig. 7. Ingestion of Congo red by plasmodium-sclerotium cells stained.
b. vacuole holding bacteria soon after ingestion.
b1. vacuole drawn later, holding clustered bacteria.
f, very fluid vacuole deformed, by pressure of the plasmodiurm.
Fig. 8. a. Ingestion of alizarin sulphate by plasmodium. Sclerotium cells stained.
m. = mucilaginous cast of vacuole immediately after discharge. (See text p. 450.)
b. Acicular crystals of alizarini sulphate as they appear before ingestion (violet), and swollen and changed by the action of the acid (red).
Fig. 9. Ingestion of alizarin sulphate by Carchesium.
a. Acicular needles without food; stored as red amorphous mass.
b. Red flakes, changing to orange and yellow (with solution) and returning to pink with re-deposition.
s. a. = small stored mass of white of egg ingested with red alizarin sulphate; colour yellow in storage.
Fig. 10. Ingestion of white of egg and calcium oxalate by Carchesium. To show persistence of calcium oxalate during digestion.
Figs. 1 and 5 in text:
'Globoids of aleurone grains. Each of these little masses of the double phosphate of calcium and magnesium is invested by a delicate, nitrogenous envelope proper to itself, and the majority (at least in the endosperm cells of Ricinus) are embedded in the proteid matter of aleurone grains (fig. 1 b, c). We have stated above that the endosperm cells are so affected by the treatment which precedes administration, that the moving substance of the plasmodium is able to break them up after ingestion, scattering even the constituents of one cell. In extreme cases this disintegrating action presses out the globoids from their normal surroundings so that they come to lie in immediate contact with the protoplasm (fig. 15). But, isolated thus or in situ they are dissolved with equal readiness (in plasmodia of like vigour), and this at a time when the colour indicators proclaim the presence of acid, and the size and form of accompanying or neighbouring aleurone grains are unaltered.' (456)
Figs. 1 and 7 in text:
'We have spoken above of potential lethargy in the initiation of actual digestion as characteristic of the Mycetozoa in a marked degree; and it may be that in this lethargy the process of aggregation is involved. At any rate a clearer conception of the possibilities of delay here is gained by study of a cuiriotis separation and distribution of large masses of inert food which occurs immediately after ingestion. When a cluster of recently enclosed aleurone grains (cp. fig. 1, b, c) or sclerotium cells is watched, one cell or grain and then another is seen to tarry, parted from the main mnass, and finally to be hurried away, it may be along some other strand of the plasmodial network. There is no such immediate scattering of living organisms after ingestion, and the nature of the force which separates inert matter thus is not obvious; we think however that the activity which manifests itself familiarly as streaming movement may exert a "kneading" pressure capable of detaching bodies as loosely bound together as are the aleurone grains of a dried endosperm cell; it may, in fact, deform a large vacuole temporarily (fig. 7, f), separate one vacuole into two, or (rarely) express a solid from the vacuole in which it lies.' (448)
Fig. 1 in text:
'it is evident that while immersion in solution (as of methylene blue, of Congo red, of litmus) is a mode of treatmient applicable to both these animals, the relatively large solid food masses with which a plasmodium can cope are uinsuited for the ingestive capacity of Carchesium, and that conversely, very finely divided matter gathered so readily into the gullet of all Vorticellidae is unsuitable for ingestion by a mycetozoon. In fact, we used milk, and the coagulation precipitate from diltute white of egg for Carchesiun; and as nutritive ingesta for the plasmodia, sclerotium cells of the sanme or different species (fig. 1, a), and the complex aleurone grains of Ricinius communis (fig. 1, b, c). The striking physical events which mark the progress of ingesta through the substance of Carchesium have recently been described at some length, and the facts of mycetozoan digestion while they harmonize in many ways with the facts of digestion as established for Amoeba, hardly go beyond them; we wish, then, to give here no more than such a summary of existing knowledge on these points as will make intelligible a subsequent discussion upon the existence and action of acid in Protozoan digestion.' (444-445)
'We chose Badhamia panicea, Lamproderma scintillans, Didymium microcarpon [note: 'In the identification of the species of these Mycetozoa we are indebted to Mr A. Lister for much kind aid.'] and other species of these genera, and of Chondrioderma, and observed small fragments of plasmodia allowed to creep in a drop of water suspended from the roof of a moist chamber.
These plasmodia (for the sake of brevity we indicate by the name any separated fragment) ingest indiscriminately if the solid matter offered is sufficiently bulky, but the exact limits which define the size of the most welcome solids are not easy to fix. Thus, we have seen 60 sclerotium cells (fig. 1, a) enclosed by a single act, and we have followed isolated yeast cells as they move in the stream of moving cell substance; the largest solid open to the attack of a plasmodial fragment, however, may naturally be expected to increase with the size of the fragnment in question, and on the other hand very small foreign bodies are often localized and traced with difficulty in the moving endoplasm.' (446)
Fig. 3 in text:
'with essential likeness in change but along slightly different lines ingesta proceed to the end of their stay within a plasmodium, and they are ejected as gelatinous "casts" of a digestive vacuole bearing any quite insoluble residue, or from a fluid vacuole after preliminary gradual loss of that fluid, or instantaneous explosive discharge (fig. 3, a, b, c) [note: 'The discharge from vacuoles was observed by Lister in the case of Badhamia utric. feeding on stereum. Journ. Linn. Soc. Vol. L. Botany.'].' (449-450)
Fig. 4 in text:
'The experiments we made under this combination of conditions were very numerous and almost uniform in result: they brought out two points with great clearness, the red colour of ingesta in the stage of storage (v. supra p. 445), the blue tint of vacuoles in periods of advanced digestion (fig. 4, d, ex). Occasionally the colouration is so faint as to be appreciable with some difficulty, and that there is never any great depth of colour is quite in harmony with the fashion of its origin, which appears to be as follows:- the vacuoles of ingestion are very faintly blue (each comprising but a small mass of fluid); during retrogression a reddish tint appears, and as the obvious vacuole is lost and each food mass is stored in turn it lies permeated by fluid of increased concentration and acid reaction (fig. 4, s). During the period of storage there is no further change, but when that re-formation of fluid which initiates the actual solution of food becomes well marked, the colour of the food mass changes, apparently paling as it passes from red to a violet tint; finally, as advanced solution frees the litmus, a blue vacuole travels to the area of discharge.' (451-452)
Figs. 5, 7 and 9 in text:
'When such ingesta as aleurone grains are watched during solution, in many experiments they swell appreciably at first and then are diminished until a small insoluble residue lies in a wellmarked vacuole; sclerotium cells (which are, indeed, more massive) never show marked swelling but they diminish in like fashion, leaving a granular resistent residue. At other times, however, enclosed food masses, notably sclerotium cells, behave in a slightly different manner; digestion proceeds as before from without inwards, but if the proteolytic action slackens or there is temporary absorption of the fluid of the vacuole, then any nodule of food as yet unchanged is invested by a homogeneous mucilaginous sphere, or in later stages, where no unaltered food is present, a viscid mould or cast of the interior of the vacuole is formed (fig. 7, mu, fig. 5, mu, fig. 9, mu, m). These appearances characterize acts of digestion in which the ingesta are somewhat massive and the solvent process is slow [note: 'In a note appended to this paper we have tabulated some more definite statements of the time relations of the digestive act; it is often much prolonged when (we may suppose) the plasmodia have suffered from confinement or the conditions of experiment. Thus, whereas we have seen the digestive change in an aleurone grain complete in 1 to 2 hours from its onset, the intermittent solution in which this mucilaginous deposit comes out most clearly may linger through 2 to 6 days.']; we ascribe them to separation of partially digested matter on loss of the vacuolar fluid, and we think that some element present is not only undissolved but insoluble, and comparable probably to that mucilaginous residue which gives so distinctive an appearance to the egesta of many Infusoria.' (449)
Fig. 5 in text:
'We have recognised in an earlier part of this paper the possibility of the very close investment of massive and inert ingesta by a plasmodium during enclosure, thus it may happen as in the case under consideration that the acid secreted later has no fluid derived from the vacuole of ingestion to which to add itself and is hardly appreciable in bulk though potent in effect. But the fluid of the digestive vacuole is clear in all cases. It is colourless at first, but the onset of solution of a food mass places a red solid in a red vacuole (fig. 5, s, d, ex); and later, the fluid of the vacuole becomes violet and then blue. The striking fluctuations in fluidity, and hence in activity of solution to which we have referred (p. 449), may coexist with variations in colour of the vacuole and its contents,- thus there may be change fromn red to blue more than once in any digestive act. A temporary suspension of digestive activity does not necessarily coincide with one phase of the colour variation, but the fluid of a vacuole disappears as a rule when its tint is violet or blue leaving a non-vacuolate solid of the same colour. In such cases there is a passing change to red before digestion recommences (fig. 5. mu), but as solution approaches completeness a steady deepening of the blue tint becomes clear, the alkalinity of a normal vacuole of ejection (if inference from the colour be justifiable) being only less than that of the surrounding water.' (454)
'The existence of violet and dark blue ingesta in Carchesium and of the black-violet tint which so soon follows enclosure by plasmodia is proof that at certain times free acid exists; the cherry red of a vacuole and its contents during solution is incompatible with the presence of free acid, but leaves it doubtful whether combinations of acid and proteid have been formed. But here the facts of change in litmus are suggestive, for we find that in the early stages of digestion a red food mass lies in a red vacuole (fig. 5, d), while later there is change to blue; at this point combinations of acid and proteid must be absent since these react with litmnus. We conclude, then, that the free acid demonstrable in the stage of storage is replaced as proteolytic activity becomnes manifest by combinations of acid with the products of this action, and that later it is not present in the vacuole even in comnbination.' (459-460)
Fig. 6 in text:
'Congo red. This substance, like alizarin sulphate, is less soluble in ditch-water than in distilled water; we have offered it to Carchesium in solution and as a gelatinous precipitate unmixed or mixed with true food-stuffs. And although in some ways a less delicately responsive reagent than others with which we have worked, its fate after ingestion supports the conclusions which we have formulated above. Free particles of Congo red are actuially almost orange-red in tint; after ingestion they usually become pink-red, violet, and lastly blue (fig. 6, s). We qualify this statement because the blue compound seems to be formed with more difficulty and to be correspondingly less stable than the products which spring from the intracellular change of litmus or even of alizarin sulphate.' (453)
'It is clear... that in the ready solution of a phosphate [note: ' Cp. p. 456. The globoids of Ricinus seeds are made up of the double phosphate of calcium and magnesium. In this connection we may state that solutions of Congo red are deepened to black-red by the passage of a stream of carbonic acid, but show no change to blue.'] we have proof, if not of its absence, at least of the presence of some other acid. We are not prepared to make further dogmatic statement, but, bearing in mind the intensity of the blue colouration whiclh may be displayed in Carchesium by one small food mass mixed with Congo red (fig. 6), and the relative sensitiveness of this reagent to organic and mineral acid, we are inclined to believe that the latter exists here.' (461)
Fig. 7 in text:
'We have looked carefully in the Mycetozoa for the aggregation which is so constant a sequel to ingestion in Carchesium [note: 'Proc. Roy. Soc. Vol. LIV. p. 468.']. But the comparative opacity of plasmodia, their fashion of ingesting and (often) the characters of their food make it especially difficult to determine that there is or is not at one point secretion of colourless viscid matter. We notice however that when carmine grains and nutritive ingesta coexist in vacuoles they cohere throughout digestion (as is the case in Amoeba), and that a vacuole of ingestion filled with moving bacteria shows them as a central cluster when they are stilled (fig. 7, b, b1). It seems probable that these appearances do indicate the binding action of retractile matter within a digestive vacuole, its deposition being less definitely localized in time than is the corresponding secretion in Carchesium.' (447-448)
'Congo red. In our experiments this substance was always given to plasmodia as a stain for nutritive matter, and we find that its intracellular history is relatively simple. The non-vacuolate ingesta darken to an opaque black-violet, and subsequent solution in vacuoles gives bright red ingesta in bright red fluid surroundings (fig. 7, ing. s. ex). From these surroundings, directly, or after there has been phasic reversion to a non-vacuolate condition and a violet colour, innutritious débris are expelled.' (456)
Figs. 8-9 in text:
'Sulphate of alizarin is only slightly soluble in the water in which Carchesium lives and thus, breaking into flocculi or forming the acicular crystals referred to above, it takes rank as finely divided matter suitable for independent ingestion, apart from its administration as an accompaniment of food. When the violet "lime crystals" (fig. 8, b) are offered alone to Carchesium they undergo some preliminary fragmentation in the wsophagus but are still recognisable as discrete spicules during progression and quiescence. Later their rigidity and individuality are lost, and the end of retrogression sees the contents of each vacuole of ingestion moulded to a red amorphous mass (fig. 9, s). When the indicator is ingested in a less alkaline form, then parallel changes are set up; the colouration passes from red through orange to yellow, a lemon-coloured vacuole with somne central insoluble residue taking the place of the loose flake which was swallowed (fig. 9, b). Alizarin sulphate is soluble in, as well as sensitive to acids: hence the formation of gelatinous ingesta from "crystals" (which have also yielded up their lime to the acid secreted round them), and hence too this developed fluidity of vacuoles when the ingested alizarin is already in the form of gelatinous flakes.' (452)
Fig. 8 in text:
'The combination of alizarin sulphate with proteid impairs the purity of its tones of colour considerably; thus stained food masses when they are ingested are usually of a reddish-violet (fig. 8, ing). They change to dull pink, then to brownish yellow, or, rarely, to pale yellow, while still non-vacuolate and in the digestive vacuoles which succeed they are pink or (very occasionally) lilac.Fluctuations in the amount of vacuolar fluid and variations in the tint of the indicator may occur, and being strictly comparable to the phases described in the career of litmusstained food need hardly be particularized further. It may suffice to say that normal egesta are pink or lilac, their colour changing or deepening to dull violet in contact with the water into which they are freed (fig. 8, m).' (455)
Fig. 9 in text:
'Concomitant ingestion of this indicator [sulphate of alizarin] and some true food-stuff introduces two variations in the aspect of ingesta;- the colours are much fainter, for but little alizarin is associated with each food mass and the yellow tint of the storage stage (a very faint, impure yellow) is more persistent (fig. 9, b, s). When this stage is over and true digestion follows, the vacuoles are faintly pink, and here as in the case of litmus the colour is so pale at times as to be almost inappreciable before the change, suggestive of decreased acidity, appears.' (452-453)
Fig. 10 in text:
'We have also watched the ingestion and fate of crystals of calcium oxalate and with the following results; mixed with sclerotium cells and thus given to plasmodia they may be found clinging to the half-digested proteid matter after 24 hours; mixed with coaagulated white of egg, and so given to Carchesium, they stand out with increasing clearness in digestive vacuoles as solution of the proteid proceeds (fig. 10, d, ex), and when ingested alone groups of them may be observed after the lapse of some hours.' (460-461)
-
CitesW.B. Hardy and L.B. Keng, 'On the Changes in the Number and Character of the Wandering Cells of the Frog induced by the presence of Urari or of Bacillus Anthracis', Journal of Physiology 15 (4) (1893), pp. 361-400.
Description:'We believe that the presence or absence of a blue colour in the vacuole depends on the predominance of one or the other of two antagonistic processes, the first setting free methylene blue as the proteid which it impregnates is dissolved, the second depositing the pigment so freed round the residual solid nidus of the vacuole. A comparable deposition of methylene blue from solution has been described in the wandering cells of the frog [note: 'W. B. Hardy and Lim Boon Keng. This Journal, XV. p. 370, 1893.']; it must not be forgotten, however, that methylene blue is not readily soluble in solutions of salts and that there is reason to believe that salts are present in a digestive vacuole. The absence of colour which is certainly striking in Carchesium may, then, have complex causes.' (458-459)
-


