Tag: haemotoxylin


|
|
|
cites |
 |
|
  |
|||
|
Explanation of Plate XI (figs. 1-15): 'Note. In the preparation of all the figures (except 15) Abbe's camera was employed and all, with the exception of two, are illustrated as they were seen with an immersion apochromatic objective (Zeiss 3 mm., 2 mm. or 1-5 mm. focus). The exceptions are Figs. 1 and 2, in the drawing of which Zeiss D. was used. Fig. 1. Section of a villus from the pyloric end of the small intestine of a guinea-pig kept on ordinary diet. Alcohol, acid ferrocyanide mixture, balsam. x 305. Fig. 2. Optical section of a slightly compressed villus from a guinea-pig after the administration of "peptonate" of iron. l, the lacteal vessel. Alcohol, ammonium sulphide, glycerine. x 305. Fig. 3. A portion of the mucosa of the intestine in a lake-lizard. e, epithelial cells, l, iron-carrying leucocytes, r, red blood corpuscles, also shown to contain inorganic iron. Alcohol, acid ferrocyanide mixture, balsam. x 620. Fig. 4. A portion of the epithelium and underlying elements of an intestinal villus of a guinea-pig after the administration of "peptonate" of iron. l, leucocyte, bc, blood capillary. Alcohol, acid ferrocyanide mixture, balsam. x 1240. Drawn with the diaphragm of Abbe's condenser removed from the microscope. Fig. 5. Portion of a section of the liver of a guinea-pig fed with "peptonate" of iron. l, leucocytes, hc, hepatic cells, bc, blood capillary. x 1240. Drawn with the diaphragm of the condenser removed. Fig. 6. A portion of the tip of an intestinal villus of a guinea-pig kept on its ordinary diet, to show the distribution in the cells of the organic iron compounds (chromatins). e, epithelial cells, l, leucocytes, a, nuclei of adenoid elements. In the cytoplasm of two of the leucocytes are found granules of an inorganic (?) iron compound. Alcohol, nitric acid alcohol, acid ferrocyanide mixture, balsam. x 1240. Drawn with the diaphragm of the condenser removed. Fig. 7. Epithelium and underlying elements from the side of a villus of a guinea-pig on the second day of the course of yolk-feeding, to show the distribution of organic iron compounds (chromatins). l, leucocytes, a, the sub-epithelial "membrane." Alcohol, nitric acid alcohol, acid ferrocyanide mixture, balsam. x 1240. Drawn with the diaphragm of the condenser removed. Fig. 8. Epithelium and underlying elements of a villus of the same animal. a, sub-epithelial "membrane," s, the secretion from epithelial cells. Corrosive sublimate, haematoxylin, eosin, balsam. x 1240. Fig. 9. Portion of a Lieberkuhnian gland of a guinea-pig to show the distribution of organic compounds of iron (chromatins) and especially of those connected with secretion. Alcohol, nitric acid alcohol, acid ferrocyanide mixture, balsam. x 620. Drawn with the diaphragm of the condenser removed. Fig. 10. Portion of a fresh villus of a guinea-pig killed five hours after being fed with stale yolk. The epithelium has been removed. Ammonium hydrogen sulphide, glycerine. x 620. Fig. 11. Intestinal epithelial cell of a villus from the same animal. Alcohol, ammonium hydrogen sulphide, glycerine. x 620. Fig. 11. Intestinal epithelial cell of a villus from the same animal. Alcohol, ammonium hydrogen sulphide, glycerine. x 820. (Zeiss oc. 4, apochr. imm. 1.5 mm.). Fig. 12. Epithelium and underlying leucocytes of a villus from the same animal, to show the absorption of the yolk chromatin. Alcohol, ammonium hydrogen sulphide, glycerine. x 620. Fig. 13. A liver cell of an Amblystoma fed artificially for four days with egg-yolk. Alcohol, sulphuric acid alcohol, acid ferrocyanide mixture, balsam. x 820. Drawn with the diaphragm of the condenser removed. Fig. 14. Yolk spherules from a hard-boiled egg, to show the distribution of the iron (of the haematogen). In the spherule on the left the elements are fewer and coarser. Sulphuric acid alcohol (for 48 hrs.), ammonium hydrogen sulphide, glycerine. x 820. (Comp. oc. 4, imm. apochr. 1.5 mm.) Fig. 15. Free border of an intestinal epithelial cell of an Amblystoma, fed with yolk. p, cell protoplasm, h, hyaline border, y, elements of yolk. Alcohol, ammonium hydrogen sulphide and glycerine (at 60° C. for eight hours). (Comp. oc. 4, imm. apochr. 1.5 mm.).' (295-297)
Fig. 4 in text: 'In very thin sections treated with the ferrocyanide mixture and mounted in balsam, the distribution of the iron was more clearly seen. Sometimes in the epithelial cells the blue reaction was a diffuse one with blue granules collected in groups here and there in the cell, in some instances it was fouind in the inner end of the cell chiefly, while again the protoplasmic processes in the hyaline border gave an intense reaction. Fig. 4 shows some of these details distinctly. In this are represented three cells, in two of which the inner ends appear loaded with iron and they were fixed in the act of transferring it to the underlying tissue.' (272)
Fig. 5 in text: 'When the dose of iron given was not great, then the iron was mainly if not wholly confined to the peripheral zone. With large doses a greater portion of the lobule was impregnated with iron, and other elements came prominently to view, especially in the "peptonate" preparations. There were leucocytes in the angles of the capillaries in all parts of the lobule but very frequently in the central portion, and their occurrence was manifested under the low power by the strong reaction which they gave for iron (fig. 5). Sometimes each cell was a mass of blue material or it contained large blue masses, in others again, the cytoplasm had a diffuse blue tint with one or more clumps of ironholding substance.' (273)
Figs. 6-8 in text: 'In the guinea-pig, as ordinarily fed, the "masked" iron exhibits in its distribution in the epithelium of the intestine very little difference from that represented in fig. 6, in which the iron is shown in the chromatin of the nuclei and in a narrow zone immediately about some of the nuclei, but in preparations from animals fed with yolk for two or three days, the epithelial cells situated on the sides of the villi and below the tips of the same have the iron distributed as represented in fig. 7, in which the whole of the protoplasm in the lower half of each cell and in the leucocytes below give a uniformly diffuse Prussian blue reaction. The epithelial cells at the tips of the villi are so much distorted by the fat present in them, that a division of each into an internal and external part is impossible, except in some cases where the absorption of fat has ceased to take place. In preparations stained with haematoxylin and eosin, the cells immediately below the tip give usually the appearance represented in fig. 8, but with this exception, that the bodies enclosed in cavities of the protoplasm in the external half of each cell shown in the figure are not present in the preparations from all the animals fed with egg-yolk.' (283-284)
Fig. 9 in text: 'Now as in the secreting cells of all sorts, and especially in those of the Lieberkühnian glands, secretory activity is associated with the presence of a chromatin in the part of the cell remote from the lumen (fig. 9), it might be urged that the increase of the "masked" iron in the inner ends of the superficial epithelial cells of a villus was due not to an iron compound absorbed, but to secretory activity bringing about an increase of the substance governing that process.' (284-285)
Fig. 11 in text: 'It may, in fact, be that the oval vesicles were not connected with absorption at all and that they were merely appearances in the rodlets, although, on this view, the green reaction in their envelopes would be difficult to explain. That the rodlets are not always simple structureless elements, I have, several times, found to be the case in preparations from the guinea-pig (fig. 11), in which each rodlet appeared to be a series of beadlets or granules.' (293)
Fig. 12 in text: 'There was evidence of the transference of yolk chromatin from the epithelial cells to the underlying elements, and this was found in those villi in which the process of fat absorption had not distorted the cells. Sections of these villi, obtained from material hardened in alcohol, when treated with ammonium hydrogen sulphide, gave preparations like that of which fig. 12 is an illustration. The inner portions of some of the cells at the extreme tip of a villus gave a faint greenish reaction immediately after the reagent was added, but in the corresponding portions of other epithelial cells the reaction was given also by granular elements lying between and among the fat droplets.' (290)
Fig. 14 in text: 'The failure of all these experiments led me to use a less abnormal kind of food; and, since, according to Miescher [note: 'Miescher. Hoppe-Seyler's med. chem. Unters. Pt. 4, p. 502, 1871.'], egg-yolk itself contains 1 to 1-5 per cent of nuclein (haematogen), that food substance appeared likely to yield the best results. I used unboiled egg-yolk, for when egg-yolk is hard-boiled the yolk spherules become thereby fixed in form, and the chromatin-holding particles are set free only when the spherules are digested, but wben the yolk is administered fresh the spherules readily undergo fragmentation and the chromatin-holding particles are liberated and put in a form in which the epithelial cells, if they possess the power, can invaginate them. In the spherules the chromatin is partly in a granular form [note: 'Miescher (loc. cit.) localised the nuclein which he discovered in egg-yolk in the granules of the yellow yolk spherules.'] and, apparently, partly as envelope material to its fat globules, the latter varying in size and shading off into the small granules in such a way as to suggest that the latter are also fat globules of almost infinitesimal size surrounded by chromatin. Fig. 14 gives a representation of two yolk spherules which were fixed by heat and in which the iron, set free by sulphuric acid alcohol, was converted by ammonium sulphide into the sulphide. In it can be seen smaller and larger fat globules surrounded by an iron-holding envelope. The fat is, therefore, closely associated with the chromatin, and as we know the former is in some way absorbed by the intestinal epithelium, the conclusion did not appear to be a strained one that both constituents are absorbed together.' (282)
Fig. 15 in text: 'With 15 mm. apochromatic immersion (Zeiss) and compensation ocular 4, the vesicles could be seen connected by a grayish line. On the protoplasmic side of the margin were also minute vesicles with greenish envelopes, apparently of the same character as those in the striated border and the reticulated protoplasm itself had a slightly greenish tinge (fig. 15).' (292)
|
||||||||
|
|
|
cites |
 |
|
|
|||
|
Explanation of Plate XXXIII (figs. 14-19): 'Fig. 14. From chamber five days in subcutaneouis tissue. Plasma-cells adhering to a cotton-fibre. Osmic vapour and carmine. Zeiss apochlr. oil and oc. 4. Fig. 15. From chamber eight days in subcutaneous tissue. Plasma-cells adhering to a hair, which had accidentally been allowed to get into the wound. Zeiss obj. D, oc. 2. Osmic acid solution and haematoxylin. Fig. 16. Inflammatory membrane from chamber eight days in the abdominal cavity; taken from a tenuous portion of the membrane. Four fibroblasts, in a film which is composed of an extremely irregularly arranged network of filaments resembling fine fibrin threads. The processes from the cell-body are continuous apparently with the fibrils of the matrix. Osmic acid vapour and haematoxylin. Zeiss apochr. oil imm. and oc. 4. Outlined with camera lucida. Fig. 17. Stellate fibroblasts and two leucocytes from same preparation as Fig. 9, more highly magnified. Zeiss apochr. oil and oc. 4. Outlined with camera lucida. Fig. 18. The modified Ziegler chamber; the sketch shows the actual size employed. Fig. 19. Portion of the chamber seen edgewise, showing the opening between the cover-glasses. Enlarged 12 times.' (576)
Figs. 14-15 n text: 'Contiguous plasma-cells or even those a little distance apart were often connected together by their processes (Figs. 1, 5, 7 and 8, Plates XXXI. and XXXII.). The bands of connection might be short thick arms or long gossamer threads of protoplasm. By similar arms and threads the cells seemed to adhere to the most diverse objects in their surrounding. The surface of the cover-glass, a filamllent of fibrin, a hair, a fibre of cotton, a lump of the cement fastening the sides of the chamber together, all afforded points to which the processes from the plasma-cells would cling (Figs. 14 and 15).' (560)
Fig. 16 in text: 'Each individual cell was of a discoid or fusiform figure, and granular, with a large clear nucleus. The edge of the disc was thin and often deeply scalloped; it merged, under all methods of staining used by us, at certain points quite imperceptibly, in a tenuous film which composed the bulk of the membrane proper. When fixed with osmic acid and after-stained with haematoxylin (Ehrlich's), this membrane is shown to contain, if not to be entirely made up of, a feltwork of filaments, like filaments of fibrin. These cross in every direction in the plane of the membrane, without prominent arrangement in any one particular sense. The individual filaments vary a good deal in size. Fig. 16, P1. XXXIII.' (561-562)
Fig. 18 in text: 'Two circular cover-glasses, each 5/8 of an in. in diameter and .006 of an in. in thickness, were fastened together so as to form a little flat glass chamber, in the manner employed by Ziegler. A strip of tinfoil placed between them at their edge along 11/12 of their circumference was cemented by shellac on each face to the corresponding surface of the cover-glass. The tiny chamber thus formed had therefore between the two ends of the strip of tin-foil an opening into the interior. The tin-foil first employed was 1/10 mm. thick; that thickness was inconvenient, as the depth of the chamber was then too great for higher powers of the microscope to explore. Tin-foil 1/20 mm. in thickness was subsequently employed. With this thickness membranes were obtained between the cover-glasses that made very satisfactory microscopical specimens. Fig. 18, Plate XXXIII.' (552-553) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
Description of figs. 1-6, Pl IV: 'Fig. 1. -Normal cornea of frog, stained with haematoxylin. Two of the branched corneal corpuscles are shown with a wandering cell, a, lying in the cell space with one of them. b b represent two of the wandering cells in the substance of the cornea; these have taken the elongated form. Fig. 2. -Normal cornea of a cat, stained with silver and carmine. The ground substance is stained brown with the silver, leaving the cell spaces unstained. In these are seen the nuclei of the branched cells stained with carmine. b b, two wandering cells in the cell spaces. Fig. 3. -Scleral edge of cat's cornea fourteen hours after central inflammation. The wandering cells, b b, are increased in number, and the communications between the spaces are larger than in No. 2. Silver and carmine. Fig. 4. -Area of general infiltration forty hours after central inflammation. The cell spaces are greatly enlarged, and broken up into small areas by the brown silver lines. The ground substance is reduced in amount, in some places represented only as small islands. Fig. 5. -Innermost limit of area of general infiltration. Here, as in No. 4, the cell spaces are greatly enlarged, and divided into small areas, in each of which the brightlystained horseshoe nucleus is seen. From this point to the centre no cellular elements are found. Silver and carmine. Fig. 6. -Two corneal corpuseles, which have taken on regenerative changes. The nuclei have increased in number, and long processes which are much branched have grown out from the parent cell.' (87)
Fig. 1 in text: 'We... find, even in the normal cornea,... [a] set of cells, which cannot be considered a part of its fixed histological elements. Their numbers are variable; in some corneas very abundant, in others few: in animals of the same species sometimes they are found in greater numbers at one portion of the tissue, sometimes at another. In fresh preparations they can be seen to pass by active amoeboid movements from one place to another, and they never, so far as we can see, stand in any fixed histological relation to the other elements of the tissue; these are the "wandering cells." Their position is not at all constant; sometimes we find them lying in the cell space along with the branched corpuscles, sometimes in the narrow communication between two spaces, sometimes as long drawn out rods in the tissue between the fibres (b, Fig. 1, P1. IV.), sometimes in the nerve lymph channels, and in one preparation I have been so forttunate as to get one seemingly in the act of passing from the nerve channel into a cell space communicating with this, half of its body lying in the channel and half in the space.' (78-79)
Fig. 2 in text: 'Proceeding now to the cat's cornea, we meet here, even in the normal state, some difference from that of the frog. The corpuscles (Fig. 2), are smaller, are more numerous, and the cell spaces communicate by larger passages than in the frog. The brightly-staining wandering cells in the normal cornea are fewer in number than in the frog's cornea, and mostly found in the cell spaces.' (83)
Fig. 3 in text: 'Let us now see what the carmine staining shows in the two parts. In the outer ring we have (Fig. 3) in each of the slightly enlarged cell spaces the large oval nucleus of the branch cell totally unchanged, and staining in all respects like the normal.' (84)
Fig. 4 in text: 'The animal is first etherized and the cornea touched with the caustic... If we examine such a cornea, say 40 hours after cauterization, and as yet unstained by carmine, the changes found can be divided into two heads. Of these the first will comprise the changes around the outer corneal edge, and the second those in the immediate neighbourhood of the eschar... In the immediate neighbourhood of the eschar the change is more pronounced, and different from anything we have hitherto seen. These changes are all the more important to us, since it is here that Stricker says the corneal corpuscles are undergoing the most rapid proliferation. In the silver preparations we see, lying in the coloured ground, groups of small white spaces with dark brown lines separating them from one another (Fig. 4); these groups correspond in shape to enlarged cell spaces.' (83-84)
Fig. 5 in text: 'Proceeding now from the corneal edge towards the eschar, we come to a region where the corneal corpuscles are wanting... the corneal corpuscles are dead-have been destroyed by the caustic. The cell spaces can be seen, most of them much shrunken, but no nucleus in them, or anything which would afford us proof of the presence of a corneal corpuscle. Lying in these cell spaces, but still more in the tissue between them, are seen multitudes of cells before described, at the scleral edge. These cells become more numerous as we proceed, until we reach a territory where the cell spaces are filled with them (Fig. 5). The spaces here are enlarged, and the communications between neighbouring ones are wider; spaces and communications are all full, and no one comparing these cells with those at the outer (i.e., the scleral) edge can doubt for a moment that they are similar.' (85)
Fig. 6 in text: 'In the corneal corpuscles which form the line outside the zone of infiltration, and which indicate the separation of the dead from the living proper corneal tissue, we find changes as early as twenty hours after cauterisation... At a later period (30 to 40 hours) the nuclei can be seen in different stages of division, and at the same time long processes are sent out from the cells into the dead tissue. These processes become longer (Fig. 6), nuclei pass from the old cell up into them, and thus they form in the dead tissue new corneal corpuscles, but n ever pus. These processes and new cells stain in all respects like parent cell from which they originated, and the nuclei have the same shape as in the old cells, though they stain more brightly, and are more granular.' (86) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
Explanation of Plate VII, figs. 1-9: 'Fig. 1. Diagram of the Cardiac apparatus and gills of Octopus, with the cardiac and respiratory nerves. The dotted lines indicate the parts covered by glands. Fig. 2. a. Muscle fibre of the ventricle of Octopus. (x 500.) Alcohol and Hematoxylin. Fig. 2. b. Portion of a muscle fibre of the ventricle of Sepiola. (x 500.) Gold chloride. Fig. 3. a. and 3. b. Plasma cells from the julletion of the kidney and auricle of Pterotrachea. (x 500.) Osmic acid and Picrocarmine. Fig. 4. Ganglion-cell from the cerebral ganglia of Pterotrachea. (x 500.) Osmic acid and Picrocarmine. Fig. 5. Plasma cell from the cutis of Pterotrachea. (x 500.) Osmic acid and Picrocarmine. Fig. 6. Connective-tissue cells; a. from the auricle, b. from the cutis of Pterotrachea. (x 500.) Osmic acid and Picrocarmine. Fig. 7. Diagram of the branchial and cardiac nerves of Aplysia. LV., RV. = Left and Right Visceral nerves. Vg. = Visceral ganglion. Gen. = Genital nerve. O. = "Olfactory organ." rn. = Left branch of the nerve to the gill. nb. = Branch to the pericardium. G. = Gill. P. = Pericardium. A. = Auricle. V. = Ventricle. a. = Aorta. Fig. 8. a. Plasma cell on auricular muscle of Helix. Fig. 8. b. Plasma cell from the connective tissue membrane between the loops of the genital duct of Helix. (Zeiss. D. Oc. 4.) Fig. 9. Diagram of the cardiac nerves of Helix. SO. = Supracesophageal ganglia. Sub. O. = Subcesophageal ganglia. Lv. = Left visceral nerve. Os. = Ovisperm duct. K. = Kidney. P. = Pericardium. A. = Auricle. V. = Ventricle. a. = Aorta.' (338-339)
Fig. 1 in text: 'The species chosen as most convenient for study was the common Poulp, Octopus vulgaris, and to it the following pages will be almost exclusively confined. The cardiac apparatus of Octopus (Fig. 1, Plate VII.) consists of a pre-branchial or venous, and a post-branchial or arterial section. The former collects the blood from the veins and drives it through the gills; the latter receives the aerated blood fromn the gills and propels it through the bodv. The former section may be divided into the following parts:- Vena Cava. Kidney Veinis. Branchial Hearts. The latter consists of :- Auricles. Ventricles. The Vena Cava (Fig. 1. VC.) is a straight thlin-walled tube, which runs down the ventral side of the liver, parallel to the intestine, towards the heart. At the level of the front end of the ventricle it divides into two branches, which being covered with glands considered to have a renal function, may be conveniently distinguished as the Kidney Veins (KV.) Each Kidney Vein, curving out towards a gill, is continued into a bulged oval body known as the Branchial Heart (BH.), their cavities being separated by a pair of valves at the junction. The Branchial Hearts have been usually held to be muscular organs devoted to propelling the blood through the gills; but that this view is not entirely correct will be shewn by the description of their minute structure given below. From each Branchial Heart a thick-walled "branchial artery" leads to the gill (G). The efferent vessels of each gill form a number of veins which soon unite to form a tuLbular auricle (LA. RA.), thin-walled but stouter than the Vena Cava. The two auricles open into the medianly situated, fleshy, somewhat globular ventricle (V), from the cavity of which the blood is prevented from returning by a pair of valves at each auriculo-ventricuilar orifice. Three efferent vessels issue from the ventricle, the most important being the great cephalic aorta (CA.), which starting from the posterior dorsal right region of the ventricle curls round to run to the head. Medianly and posteriorly is given off a small genital artery (G a.); and from the anterior ventral edge a small bulb gives rise laterally to a fine artery to each auricle and to a somewlhat larger visceral artery (V a.) medianly to the intestine.' (263-264) 'The whole cardiac system is supplied by a pair of nerves from the pleural ganglia in thie head... Their relations to the neighbouring organs and directions for finding them have been accurately given by Fredericq... Their branches and ganglia need however a more detailed description (Fig. 1). On issuing from the skull, each nerve appears as a double cord from which various small branches are given off. The cords on either side end in a small ganglion lying under the liver, from which go nerves to the body wall and the columnar muscle inserted into the mantle, while a single main trunk is continued downwards towards the heart. Just in front of the auricle it dilates into a ganglion, which may be called the 1st Cardiac Ganglion (C). From this issue a fine nerve to the generative duct, a nerve which enters the auricle, gives off a branch there and then passes through to the ventricle, and lastly a stout nerve which runs dorsally to the auricle down to the branchial heart, where it is connected with a ganglion-the 2nd Cardiac Ganglion (Cr). From this ganglion go nerves to the substance of the branchial heart and a short way into the kidney vein, but the main trunk proceeds to the gill, at the base of which it expands into the Branchial Ganglion (B). No other ganglia are revealed by dissection on the visceral nerves or their branches... From the pleural ganglia there also runs on either side a stout nerve ending in tlle stellate ganglion (s) and containing the motor fibres for the mantle; so that the gills and heart are connected by nervous structures with the motor organs of respiration.' (267-268)
Fig. 2 in text: 'The fibres of the ventricle are of an elongated fusiform shape, with long tails, but are shorter and thicker than those of the auricle. Like the others they have no sarcolemma. Thus far they resemble the ordinary smooth muscle cells of vertebrata, but in addition they possess a fine but regular transverse striation very like that of the branchial heart, the fibres of which they closely resemble. Careful focussing also shows in the fibre indications of a granular core of a different nature to the outer zone. Both of these features are best seen in osmic acid or alcohol preparations (Fig. 2a) [note: This axial column of apparently less differentiated substance has been noticed by most observers of molluscan muscles. Thus Ranvier (Traite technique d'Histologie, p. 851), figures a gold chloride preparation of the retractor muscle of Helix pomatia showing distinctly the "cordon protoplasmique"; Leydig and Kölliker saw it in the buccal muscles of Gasteropoda, and H. Muller and F. Boll in the branchial hearts of Cephalopoda. (See F. Boll, Arch. f. mikr. Anat. Bd. v. Suppl. Hft. p. 28).'].' (266) 'while the ventricular fibres of Eledone resemble those of Octopus, those of Sepia present a bolder striation, and in Sepiola the difference is still greater. In this animal the fibres are much larger, and the average distance between the striations is 3.3m.; the dark bands are very sharp, but narrow; and the margin of the fibre is bulged out opposite each broad clear disc (Fig. 2. b). The central core is much more distinct than in Octopus, occupying about one third of the diameter of the fibre, and staining very deeply with gold chloride, while the outer zone remains clear. It also shows a beaded outline corresponding with the striations. [note: 'Transverse striation has also been noticed - chiefly in hearts or the buccal mass - by many observers, a list of whom is given by Boll (loc. cit.) and by F. Darwin (Journ. of Anat. a. Phys. Vol. x. Part Ii. April 1876, p. 506). Dogiel (Arch. f. mikr. Anat. Bd. 14) has also seen it in various hearts, (Pecten, Anodon, Aplysia, Helix) and Haller (Gegenbaur's Jahrbuch, 1883) in the hearts of Fissurella and Haliotis. Margo (Wien. Sitzusngsb, Bd. 39) has described the black bands in the striated shell muscle of Anodon as doubly refracting.'].' (266 [note 266-267])
Figs. 3-5 in text: 'Two kinds of elements however occur [in Pterotrachea] which might be mistaken for nerve cells, and which I believe have been taken for such in other Molluscs. The first of these are large, moderately granular, often roundish or oval cells, with a not very distinct oval nucleus placed near the periphery of the cell, and in which a nucleolus is sometimes visible. A distinct capsule is not present. These cells occur in great abundance on the walls of the rhythmically contractile kidney, and a few are usually to be found in the auricle [note: 'Although in teased preparations of auricle these cells were often found, I am inclined to doubt whether they occur normally in the auricle itself, and to think that in these cases they have appeared from adhering fragments of the closely connected kidney, which owing to the transparency of the tissues it is difficult to cut clearly off at the boundary. In preparations where the auricle was cut through at a distance from the kidney no such cells were found. At the same time it is possible that they may occasionally wander from the kidney to the auricle. They are also found in the pericardial wall.']. I have not observed them in the ventricle. Although the approximately oval form (Fig. 3 (a) Pl.VII.) is perhaps the commonest shape, yet it is by no means constant. In many cells short blunt processes or pseudo-podia are seen (b) which suggest the power of amoeboid movement, and occasionally a single such process may occur, causing some resemblance to a ganglion cell with the stump of a nerve fibre. The number and variations of the processes however oppose this idea, and a comparison of such a cell with a true ganglion cell (Fig. 4) at once shews a number of differences in appearance. Further these cells are never found in connection by fibres with either nerve or muscle, and are shewn to be very loosely applied to the tissues by the fact that in teased preparations large numbers of them become entirely detached and float like the blood corpuscles in the fluid in which they are mounted. Lastly, they occur not only in the parts mentioned, but in the connective tissue all over the body, as is well seen in preparations of the cutis, where they (Fig. 5) abound. These facts point to the conclusion that they are wandering connective tissue cells; and we may almost certainly identify them with the "plasma cells" which have been shewn by Brock [note: 'J. Brock. " Untersuch. ii. d. interstitiellen Bindesubstafiz der Mollusken." Zeit. f. Wios. Zool. Bd. 39. 1883.'] to be so characteristic of the connective tissue of some other Molluscs (Aplysiadae, Pulmonatae).' (321-322)
Fig. 6 in text: 'The second form of element which might be considered nervous [in Pterotrachea] is of much smaller size and more delicate structure. These cells again I found only where connective tissue was abundant. They are granular branched cells with a usually indistinct nucleus. Occasionally the branches may be reduced to two, and there then appears some resemblance to a small bipolar ganglion cell (Fig. 6 (a)). These cells however are nothing more than the ordinary connective tissue cells, which give off processes to form fibres and are found everywhere throughout the connective tissue (Fig. 6 (b)). A comparison of the two figures makes plain the identity of those of the heart with those of the cutis. These also are described and figured by Brock in the connective tissue of Aplysia as stellate connective tissue cells, and very similar cells by Haller as occurring 'in small quantity in the auricle of Fissurella and as forming a second type of ganglion cell.' (323)
Fig. 7 in text: 'Apart from the improbability of the existence of apolar ganglion-cells in the heart, the incorrectness of Dogiel's drawing of the heart and the adjacent nerves causes great doubt as to the accuracy of his interpretation of these cells. In his figure (Taf. V. a. Fig. 15) of the "branchial" ("visceral," Spengel) ganalion he entirely overlooks the olfactory organ to which the right nerve runs, and he represents the left nerve as of equal thickness and as running straight to the auricle. But instead of such a short thick left nerve, there come off from the left half of the double "visceral " ganglion two distinct long nerves. Of these one supplies the generative duct, while the other runs on to the gill. Here it divides into two, the left one running on straight towards the hind end of the gill while the right curls round anteriorly and appears to end in the pericardium near the origin of the auricle (Fig. 7).' (325)
Fig. 8 in text: 'In the auricle of the Snail may be found upon the muscle bundles [note: 'Both the auricle and ventricle are formed of a meshwork of bundles of striated muscle fibres, that of the ventricle being denser and thicker.'] a number of plasma cells which are, there is little doubt, identical with Dogiel's "apolar!" ganglion cells. Some of these cells of an oval or pear-shaped form present a considerable resemblance to ganglion cells, and if only these were observed might be considered such. But when others of varying shapes, and others in process of division are seen, such an idea becomes untenable; and when the striking similarity of these cells with the plasma cells of the connective tissue is noticed, no doubt can remain of the identity of the two (Fig. 8).' The various stages of division also which are met with, from a group of two or three contiguous cells to a nest of closely packed small ones, point clearly to the true nature of these elements. True ganglion cells I believe do not exist in the Snail's heart.' (327)
Fig. 9 in text: 'The heart... receives nerves both at the auricular and aortic ends [note: 'A similar double nerve supply has been described by Haller, loc. cit. as existing in Muricidae and Fissurella.']. (Fig. 9.) The discovery of this nerve to the heart at once necessitated a reconsideration of the results obtained by previous workers, and the question of its function became important.' (328) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
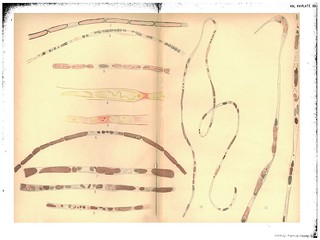
Explanation of Plate XII (figs. 1-11): 'The figures with one exception were drawn under a zeiss D and ocular 4. The outlines were filled in with the Camera Lucida and an attempt made to colour the drawings like the particular specimen under examination. Fig. 1. To show the first segmentation of the myeline in degeneration. Each segment completely enclosed in myeline, with a central piece of the axis cylinder. Experiment XX. 2 days. R. S. middle of forearm. Osmic acid and haematoxylin. Fig. 2. To show the secondary fragmentation of the myeline (extreme case). Exp. XX. 2 days. L S. middle of forearm. Osmic acid and haematoxylin. Fig. 3. To show the remnants of the disintegrating axis in the segments of myeline. Exp. XIII. 4 days. R. S. 3 in. below wound. Gold and haematoxylin. Nucleus stained blue in haematoxylin. Fig. 4. The same. Exp. IIl. 7 days. R. S. Below wound 2 in. Picric acid and fuchsin stain. Fig. 5. The same. Exp. III. 7 days. R. S. Below wound 2 in. Picric acid and fuchsin stain. Fig. 6. To show the more rapid fragmentation of the myeline in the neighbourhood of the nuclei. Exp. III. 7 days. Gold and haematoxylin. Nucleus stained blue. Fig. 7. To show the multiplication and migration of the nuclei of the sheath and greater absorption near nuclei. Exp. III. 7 days. Gold anld haematoxylin. Fig. 8. The same. Exp. III. 7 days. Gold and haematoxylin. Fig. 9. An instance of apparent indirect division of nucleus. The progressing fragmentation of myeline. Exp. III. 7 days. Gold and haematoxylin. Fig. 10. (Drawn under obj. C. oc. 4). To show fragmentation of myeline especially at nuclei, and the material containing fragments of myeline which fills the fibre. Exp. III. 7 days. Gold and haematoxylin. Fig. 11. To show progressive absorptionl of myeline; the proliferation of the nuclei; the position of the nuclei with reference to the balls of myeline; and the collapse of the empty sheath. Exp. XI. 9 days. R. S. middle of forearm. Gold and haematoxylin.' (402)
Fig. 1 in text: 'The segments first formed in degeneration, unlike the normal appearance of the segments of Lantermann seen in fibres after the action of osmic acid, have a complete envelope of the myeline, as shown in Fig. 1, and the small interspaces between them are filled with a colourless material. In each of these segments is a piece of the original axis cylinder, at this time apparently unchanged. Many of the older histologists have asserted that the axis remains unaffected during the degeneration. On the contrary, even at this early stage the breaking of the myeline into segments is accompanied by, or causes a simultaneous breaking of the axis.' (371)
Figs. 2-8 in text: 'Shortly after the first cylindrical segments of myeline and axis are formed an irreaular fragmentation occurs in these segments in most parts of the fibre. The segmetnts break up into smaller or larger irregular pieces or balls, an extreme case of which is shown in Fig. 2. Very frequently in these irregular masses of myeline remnants of the axis cylinder can be clearly distinguished in all stages of disintegration as shown in Figs. 3, 4, 5. It should be stated that some of the large segments frequently persist in different portions of a fibre long after tlle other segments have broken into small pieces and become partially absorbed,-the process is very irregular. It is at the time that this breaking up of the cylindrical segments into smaller fragments becomes apparent that the increase in the size of the nuclei of the sheath and the growth of the protoplasm surrounding these rnuclei become clearly marked. In fact this secondary fragmentation is always visible first in the neighbourhood of the nuclei as shown in Figs. 6, 7, 8. The large rounded nuclei lie in the middle of the fibre, and close to them are the small drops or balls of myeline. For this reason we believe that one cause of this secondary fragmentation is to be found in the absorption which takes place under the influence of the nucleus and its suirrounding protoplasm.' (372)
Fig. 6 in text: 'As evidence of the actual absorption which is beginning to take place one finds, at this time, say the seventh day, in the fibres stained with osmic acid or gold, some of the balls of myeline in the neighbourhood of the nuclei left colourless by the stain, see Fig. 6. In the later stages the same fact may frequently be noticed at different parts of the fibre and always most clearly near the nuclei. See Fig. 11.' (372-373)
Figs. 7-8 in text: 'An interesting fact in connection with the inultiplication of the nuclei is the way in which they migrate. In the beginning, of course, there is a single nucleus to each internode. At the time the secondary fragmentation of the myeline is fully under way one frequently finds a number of nuclei in the space which an internode would occupy. Sometimes they are in pairs as though from a recent division, but in other cases one or more large masses of myeline will be found between. See Figs. 7 and 8. This latter appearance has been used to support the theory that there are several nuclei present in the internode in a normal fibre, but that they are hidden by the myeline. Such a view as this it is not necessary to consider at present. The only explanation of the appearance described that seems reasonable is that after division the nuclei migrate or may migrate to some distance and start the process of absorption at a new place.' (373)
Fig. 9 in text: 'By the 7th day a very active proliferation of the nuclei of the sheath has begun. The increase in their number is very striking. Our methods of hardening were not such as to show the method of multiplication of the nuclei, though we often found nuclei showing a dumb-bell form, i.e., an elongated nucleus constricted in the middle as though multiplying by direct division. There can be little doubt that the division is indirect, as we sometimes found, even after hardening in Mueller's solution, nuclei like that pictured in Fig. 9 which evidently represents a badly preserved mitosis.' (
Figs. 10-11 in text: 'From the 7th day to the 14th day the process of absorption of the balls of myeline with their contained fragments of axis cylinder goes on actively, yet quite irregularly. Fig. 10 from a nerve after 7 days shows very well the breaking-up of the myeline and the formation in between, especially at the nuclei, of an apparently liquid substance in which are contained numerous fragments of the old myeline. As the absorption progresses and the fragments of myeline become smaller and less numerous the direct participation of the nuclei in the process becomes more evident. The nuclei are much more numerous and are found clustered in and about the remaining balls of myeline as shown in Figs. 11 and 12, representing a degeneration of 9 and 14 days respectively. One often sees bits of the myeline partially imbedded in a nucleus, and this appearance is found from this time on well into the later stages of regeneration, as long, in fact, as any of the myeline remiains unabsorbed. After 14 days absorption has gone so far that long stretches may be found, as shown in Figs. 12 and 13, in which only small fragments of myeline are present. At such places the fibre consists of a homogeneous, apparently liquid substance lying in the old sheath, and of many nuclei, often in pairs or groups, the latter giving indication of an active proliferation. Yet at this time, 14 days, and even later, one sees many fibres in which the absorption has lagged behind the condition of what may be considered a typical fibre of this period. In one and the same fibre places will be found in which absorption has made rapid progress in spots, all the myeline having disappeared, while in other spots the large cylindrical segments have suffered scarcely any change. Examples of this are pictured in Figs. 14 and 15. However irregularly the process may go on the final outcome is the complete absorption of the remnanits of the old myeline and axis; though as we have said before, balls of the myeline may be found in certain fibres long after this period, even at the time when fully formed new fibres have beeu produced. As the absorption proceeds the old sheath collapses more or less. It seems at first to contain a liquid material with some debris of the old myeline, but this too finally disappears and the beginning of the actual process of regeneration is inaugurated by the formation of new protoplasmic material around the numerous nuclei contained in the fibres.' (373-374) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
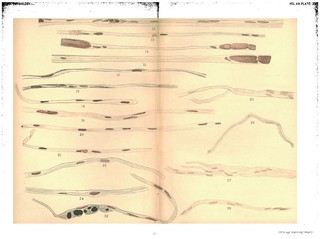
Explanation of Plate XIII, figs. 12-28: 'Fig. 12. To show still further absorption of myeline, the increase in nuclei and the way they are grouped round the balls of myeline. The collapsed tube between the nuclei. A bit of endoneural sheath lying at the side of the fibre. Exp. VII. 14 days. R. S. Gold and haernatoxylin. Fig. 13. To show the nearly complete absorption of myeline and the nuclei lying free in the tubes. Exp. VII. 14 days. R. S. Gold and haematoxylin. Fig. 14. To show the irregularity of absorption. A portion of the tube free from myeline next to a portion still showing large segments. Exp. VII. 14 days. L. S. Gold and haematoxylin. Fig. 15. The same. Exp. VII. 14 days. R. S. Gold and haematoxylin. Fig. 16. To show the beginning of the formation of an embryonic fibre, the growth of protoplasm at the nucleus. Exp. XIV. 3 weeks. R. S. 3 in. below wound. Gold and haematoxylin. Fig. 17. The same. Later stage. Exp. XIV. 3 weeks. L. S. 3 in. below wound. Gold and haematoxylin. Figs. 18, 19, 20. To show stages in the formation of the embryonic fibres, 18 from Exp. IX. R. S. (3 weeks), 19 from Exp. IX. R. S. (3 weeks) and 20 from Exp. VIII. R. S. (3 weeks). Gold and haematoxylin. Fig. 21. To show the flattened shape of the embryonic fibres often seen in the preserved specimens. Exp. VIII. 3 weeks. L. S. Below wound 3 in. Gold and haematoxylin. Fig. 22. To show the large balls of myeline in the plasma cells lying between the fibres. Exp. XVII. R. S. 4 weeks. 4 in. below wound. Osmic acid and haematoxylin. Figs. 23, 24. To show fully formed embryonic fibres and appearance of new sheath. Exp. XXVIII. 3 weeks. Br. to flex.-profundus. Osmic acid and haematoxylin. Figs. 25, 26, 27, 28. From the peripheral end of human ulnar 6 1/2 months after section, union with central end not made. To show formation of "embryonic fibres." Gold and haematoxylin.' (402-403)
Figs. 12-15 in text: 'From the 7th day to the 14th day the process of absorption of the balls of myeline with their contained fragments of axis cylinder goes on actively, yet quite irregularly. Fig. 10 from a nerve after 7 days shows very well the breaking-up of the myeline and the formation in between, especially at the nuclei, of an apparently liquid substance in which are contained numerous fragments of the old myeline. As the absorption progresses and the fragments of myeline become smaller and less numerous the direct participation of the nuclei in the process becomes more evident. The nuclei are much more numerous and are found clustered in and about the remaining balls of myeline as shown in Figs. 11 and 12, representing a degeneration of 9 and 14 days respectively. One often sees bits of the myeline partially imbedded in a nucleus, and this appearance is found from this time on well into the later stages of regeneration, as long, in fact, as any of the myeline remiains unabsorbed. After 14 days absorption has gone so far that long stretches may be found, as shown in Figs. 12 and 13, in which only small fragments of myeline are present. At such places the fibre consists of a homogeneous, apparently liquid substance lying in the old sheath, and of many nuclei, often in pairs or groups, the latter giving indication of an active proliferation. Yet at this time, 14 days, and even later, one sees many fibres in which the absorption has lagged behind the condition of what may be considered a typical fibre of this period. In one and the same fibre places will be found in which absorption has made rapid progress in spots, all the myeline having disappeared, while in other spots the large cylindrical segments have suffered scarcely any change. Examples of this are pictured in Figs. 14 and 15. However irregularly the process may go on the final outcome is the complete absorption of the remnanits of the old myeline and axis; though as we have said before, balls of the myeline may be found in certain fibres long after this period, even at the time when fully formed new fibres have beeu produced. As the absorption proceeds the old sheath collapses more or less. It seems at first to contain a liquid material with some debris of the old myeline, but this too finally disappears and the beginning of the actual process of regeneration is inaugurated by the formation of new protoplasmic material around the numerous nuclei contained in the fibres.' (373-374)
Figs. 16-24 in text: 'Regeneration in the Peripheral End:- The increase of protoplasm round the nuclei is shown in its different stages in Figs. 16, 17, 18. At first, where the nuclei lie singly, the new protoplasmic formation gives the appearance of spindle-shaped cells or fibres such as have often been described as taking an active part in the formation of the new fibres, - Fig. 16. Later the increased formation of new material results in filling the old sheath with a continuous band or fibre of protoplasm in which the nuclei are imbedded, as shown in Figs. 19, 20, 21, 22, 23, 24. The fibre as now formed represents the completion of the first stage in regeneration.' (374)
Fig. 21 in text: 'The embryonic fibres, it should have beeni stated, often appear in the hardened specimens as flattened ribbons, instead of cylindrical threads, as shown in Fig. 21.' (375)
Figs. 23-24 in text: 'After the formation of the complete embryonic fibre a new sheath is made by a differentiation of the peripheral layer of the protoplasmic thread. The appearances which have led us to this view are shown in Figs. 23 and 24, in which the newly-formed fibre with its sheath lies within the old sheath. The old sheath is displaced outward and one may suppose that it eventually forms a part of the endoneural connective tissue which is at this time abtundant.' (375)
Fig. 23 in text: 'there is a strong probability that many of the old fibres do not succeed in regenerating, but go to ground completely. One often meets specimens such as are represented in Fig. 23, in which large balls of myeline are found on the sides of new fibres, engulfed in the cytoplasm of the wandering cells.' (375-376)
Fig. 24 in text: 'specimens like that represented in Fig. 24 may explain what Schiff and others have mistaken for a persistent axis cylinder. Certainly our modern methods of staining prove that after the first week or ten days there are no distinguishable fragments of the old axes remaining in the fibres of the peripheral end of the nerve.' (377)
Figs. 25-28 in text: 'Our longest experiments upon a nerve cut but not sutured, and in which reunion was prevented by removing a piece of suitable length, were experiment XXVI., upon a rabbit examined 70 days after the operation, and XXII., central ulnar of dog, 75 days. In addition, however, we have had the opportunity of examining a portion of the peripheral end of a human ulnar nerve which had been cut accidentally six and a-half months previously. We owe the opportunity to the kindness of Dr T. A. Mc Graw, who in an operation for secondary suiture removed small portions of the central and peripheral ends and sent them to us in Mueller's solution. Figs. 25, 26, 27, 28 are from drawings of specirmens prepared from the peripheral end of this nerve after treatment with the potash gold stain of Freud and Böhmer's haematoxylin. They show that regeneration had advanced to the stage of the formation of embryonxic fibres, but that there were no signs of an axis cylinder or myeline sheath. Similar preparations were obtained from the animals upon which section without suture was made. Cross sections of the peripheral end of the human nerve are shown in Fig. 29, stained with gold and haematoxylin and in Fig. 30, stained in haematoxylin alone. The drawing in Fig. 29 was made from a portion of the nerve in which some of the nuclei show a central nucleolus in cross section, simulating the appearance of. new nerve fibres with a central axis. This is better shown in Fig. 31, taken from the ulnar nerve of a dog after sutture but before regeneration.' (376-377) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
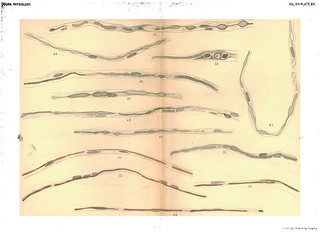
Explanation of Plate XIV (figs. 29-42): 'Fig. 29. Cross-section of human ulnar peripheral end 6 1/2 months after section, union with central end not made, To show cross-section of bundles of embryonic fibres and the appearance of .the nuclei (only one bundle filled in). Gold and haematoxylin. Fig. 30. The same. Stained in haematoxylin alone. Fig. 31. Cross-section of pelipheral ulnar 4 weeks after division. After suture but before complete regeneration. To show the resemblance of the nuclei and their nucleoli to cross-sections of young nerve fibres. Exp. XVII. 4 weeks. L. S. Gold and haematoxylin. Figs. 32, 33. Embryonic fibres in peripheral ulnar of rabbit, 7 weeks after section without union to central end. To show apparent attempt at the formation of myeline. Exp. XXVI. Osmic acid and haematoxylin. Figs. 34, 35. To show the apparent beginning of a myeline transformation in the protoplasm of the embryonic fibres. Exp. XXVIII. 3 weeks. From branch to flexor profundus. Osmic acid and haematoxylin. Figs. 36, 37. The first, to show the isolated drops of newly-formed myeline, the second, to show the varicose tube formed by their union. (Portions of the same fibre.) Exp. XXVIII. 3 weeks. Just below wound. Gold and haematoxylin. Fig. 38. To show the isolated drops of newly-formed myeline and the processes which unite them. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin. Figs. 39 and 40. To show the same and the staining of the balls of myeline in the haematoxylin. Each of the fibres shows a few fragments of old myeline unabsorbed lying near the nuclei. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin. Figs. 41 and 41a. The same. To show discontinuous formation of the myeline tube. Exp. XXVIII. Below wound. Osmic acid and haematoxylin. Fig. 42. The same. Exp. XXVIII. Below wound. Osmic acid and haematoxylin.' (403-404)
Figs. 29-31 in text: 'Our longest experiments upon a nerve cut but not sutured, and in which reunion was prevented by removing a piece of suitable length, were experiment XXVI., upon a rabbit examined 70 days after the operation, and XXII., central ulnar of dog, 75 days. In addition, however, we have had the opportunity of examining a portion of the peripheral end of a human ulnar nerve which had been cut accidentally six and a-half months previously. We owe the opportunity to the kindness of Dr T. A. Mc Graw, who in an operation for secondary suiture removed small portions of the central and peripheral ends and sent them to us in Mueller's solution. Figs. 25, 26, 27, 28 are from drawings of specirmens prepared from the peripheral end of this nerve after treatment with the potash gold stain of Freud and Böhmer's haematoxylin. They show that regeneration had advanced to the stage of the formation of embryonxic fibres, but that there were no signs of an axis cylinder or myeline sheath. Similar preparations were obtained from the animals upon which section without suture was made. Cross sections of the peripheral end of the human nerve are shown in Fig. 29, stained with gold and haematoxylin and in Fig. 30, stained in haematoxylin alone. The drawing in Fig. 29 was made from a portion of the nerve in which some of the nuclei show a central nucleolus in cross section, simulating the appearance of. new nerve fibres with a central axis. This is better shown in Fig. 31, taken from the ulnar nerve of a dog after sutture but before regeneration.' (376-377)
Figs. 35-41 in text: 'Whatever theory of the immediate cause of the formation of the myeline may be the true one, there can be no doubt that it is first found as disconnected drops. These may afterwards become united by slender processes to form a bead-like string which sooner or later elongates to an even tube, or the drops mnay first elongate to form cylindrical segments which eventually unite to form continuous, delicate tubes of myeline. Both of these processes, with the intermediate stages, are shown in Figs. 36 to 55 better than we could describe them in words. What becomes of the numerous nuclei scattered along the embryonic fibre it is not possible to say, other than that they disappear. In some cases, as in Figs. 38, 39, 42, they disappear rapidly as the myeline tube is formed, while in other cases they persist for a much longer time, the newly-formed myeline tube winding in and out among them in a very beautiful manner (see Figs. 37, 51, 55). We can only suppose that they disappear by absorption, as their nutritive relations with reference to the protoplasmic contents of the fibre become less and less important. With reference to the nodes and internodes of Ranvier, it is evident that no simple hypothesis, such as the development of each internode from a single cell, will fit the facts as they appear in regenerating fibres. The developing internodes and nodes are plainly shown in Figs. 51 to 55, but why the ends of the internodal tubes do not fuse together is difficult to explain.' (379-380)
Figs. 36-41 in text: 'After the formation of the embryonic fibres the additional steps necessary for a complete regeneration consist in the production of a new myeline sheath and a new axis cylinder... Figs. 36, 37, 39, 40 and 41 give an idea of how the myeline first appears. As shown in these figures, it appears first as irregular deposits in the protoplasm of the embryonic fibres, and usually first in the immediate neighbourhood of the nuclei. Delicate prolongations of the myeline are often seen running from one small mass of the myeline to another, and eventually these latter become connected together, forming a varicose tube, shown in various stages in Figs. 36 to 51. With reference to the immediate origin of these deposits, we must confess ourselves in doubt. In many cases we obtained specimens such as are shown in Figs. 34 and 35, in which clear bead-like spaces are seen forming in the protoplasm. The general arrangement of these spaces strongly suggests that they mark the beginning of the transformation of the protoplasm to myeline; a step farther and the chemical change will be such as to give the usual staining with osmic acid or gold as seen in Figs. 36-38, etc. On the other hand, the first deposits of myeline, especially when stained in osmic acid, often bear a striking resemblance to the nuclei, both in shape and in the fact that they show a rather distinct staining with the haematoxylin. (See Figs. 38, 39, 40, 42, etc.) There is no doubt that as the myeline forms many of the numerous nuclei scattered along the fibre disappear.' (378-379) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
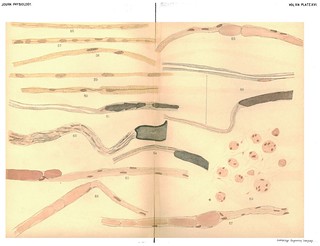
Explanation of Plate XV (figs. 42-55): 'Fig, 43. To show varicose appearance of newly-formed tube of myeline. Exp, XXVIII. 3/4 in. below wound. Osmic acid and haematoxylin. Fig. 44. The same. Exp. XXII. R. S. near wrist. Osmic acid and haematoxylin. Fig. 46. To show the formation of the myeline tube without varicosities. Exp. XXVIII. 3/4 in. below wound. Osmnic acid and haematoxylin. Fig. 47. To show union of isolated drops of myeline to form a tube. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 48. To show discontinuous formation of myeline sheath. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 49, The same. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 50. To show the persistence of balls of old myeline as yet unabsorbed and their position with reference to the nuclei. Exp. XXVIII. Osmic acid and haematoxylin. Nuclei stained in haematoxylin. Fig. 51. To show the formation of a node of Ranvier. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 52. To show the newly-formed myeline tube lying in the embryonic fibre. Exp. IX. 3 weeks. From dorsal cutaneous branch. Gold and haematoxylin. Fig. 53. The same. Exp. IX. L. S. Dorsal cutaneous branch. Gold and haematoxylin. Fig. 54. To show the formation of nodes of Ranvier. Exp. IX. L. S. Dorsal cutaneous branch. Gold and haematoxylin. Fig. 55. To sbow newly-formed myeline tube with node of Ranvier and persistent nuclei. Exp. IX. L. S. 3 weeks. At wound. Gold and haematoxylin.' (404-405)
Figs. 42-55 in text: 'Whatever theory of the immediate cause of the formation of the myeline may be the true one, there can be no doubt that it is first found as disconnected drops. These may afterwards become united by slender processes to form a bead-like string which sooner or later elongates to an even tube, or the drops mnay first elongate to form cylindrical segments which eventually unite to form continuous, delicate tubes of myeline. Both of these processes, with the intermediate stages, are shown in Figs. 36 to 55 better than we could describe them in words. What becomes of the numerous nuclei scattered along the embryonic fibre it is not possible to say, other than that they disappear. In some cases, as in Figs. 38, 39, 42, they disappear rapidly as the myeline tube is formed, while in other cases they persist for a much longer time, the newly-formed myeline tube winding in and out among them in a very beautiful manner (see Figs. 37, 51, 55). We can only suppose that they disappear by absorption, as their nutritive relations with reference to the protoplasmic contents of the fibre become less and less important. With reference to the nodes and internodes of Ranvier, it is evident that no simple hypothesis, such as the development of each internode from a single cell, will fit the facts as they appear in regenerating fibres. The developing internodes and nodes are plainly shown in Figs. 51 to 55, but why the ends of the internodal tubes do not fuse together is difficult to explain.' (379-380)
Fig. 42 in text: 'the first deposits of myeline, especially when stained in osmic acid, often bear a striking resemblance to the nuclei, both in shape and in the fact that they show a rather distinct staining with the haematoxylin. (See Figs. 38, 39, 40, 42, etc.) There is no doubt that as the myeline forms many of the numerous nuclei scattered along the fibre disappear.' (379) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
Explanation of Plate XVI (figs. 56-69): Figs. 56, 57, 58, 59. From nerve stained in picric acid atid haematoxylin. Exp. XIX. 7 weeks. R. S. J in. below wound. 59. An embryonic fibre. 58. Newly-formed fibres with myeline and axis. 57 and 56. Newly forming fibres, myeline as a varicose tube. Axis also present but not stained in the interior of the swellings of myeline. Shows how quickly after the formation of the myeline the axis grows down from above. Fig. 60. To show the branching of the axis cylinder where an old fibre passes into several new fibres, central end of ulnar. Exp. XXlI. Cut for 75 days and union with peripheral end prevented. Picric acid and haematoxylin. Fig. 61. To show what seems to be the outgrowth of an axis cylinder from an old fibre toward a new one. New myeline lies in the embryonic fibre as a continuous delicate sheath. Exp. XVII. 4 weeks. R. S. Gold and haematoxyiin. Fig. 62. To show the formation of several new fibres in the degenerated portion of a single old fibre at the central end of wound. Exp. XVII. 4 weeks. L. S. wound of median ulnar suture. Osmic acid and haematoxylin. Fig. 63. The sanme. Human ulnar nerve. Central end of nerve 6 1/2 months after injury. No union with peripheral end. Gold and haematoxylin. Fig. 64. To show junction of old with new fibre at the central end. Exp. XVIII. R. S. wound. Osmic acid and haematoxylin. Fig. 65. The same. Human ulnar. Central end 6 1/2 months after section and no union with peripheral end. The continuation of the protoplasm of the "embryonic fibre" into the hypertrophied protoplasm surrounding the nuclei of the old fibre, and the fragmentation of the end of the old myeline to be specially noted. Gold and haematoxylin. Fig. 66. To show junction of old and new fibres, and the new fibre with its thin myeline sheath lying in the protoplasm of the "embryonic fibre." Exp. XVII. R. S. wound. Gold and haematoxylin. Fig. 67. To show junction of old and new fibres. Central end of human ulnar, 6 1/2 months after injury and no union with peripheral end. The fragmentation of the myeline and the hypertrophy of the protoplasm round the nuclei of the old fibre to be noted. Gold and haematoxylin. Fig. 68. The same. Fig. 69. Cross-section through the bulbous enlargement of central end of human ulnar, 6 1/2 months after injury and no union with central end. Shows the increase in the endoneural connective tissue as well as in the nerve fibres. The fibre marked + shows cross section of new fibres in same sheath with old. Compare with figs. 63, 65.' (405-406)
Figs. 56-57 in text: 'when the new myeline is evidently just formed no definite axis can be demonstrated by the gold stain. With the picric acid and haematoxylin stain, on the contrary, the newly formed axis is clearly seen, even at that early stage in the formation of the myeline tube when it exists as a string of bead-like swellings (see Figs. 56 and 57). It follows then that though the myeline sheath probably begins to form before the axis cylinder can be distinguished, the latter appears shortly afterward, before the new fibre has gone far in its development.' (380-381)
Fig. 61 in text: 'Very many apparent examples of this branching of the axis were found in this specimen, but owing, to the intricate way in which the fibres were twisted and the possibilities of deception arising therefrom, the connection of the old and new axes could not be satisfactorily followed. The example given in the drawing, and some others, were, however, quite distinct and seem to us to give fairly satisfactory histological proof that in regeneration the new axis cylinders are outgrowths from the axes of the uninjured fibres of the cenitral end. Fig. 61 gives an apparent example from another experiment of the outgrowth of the axis.' (382)
Figs. 62-69 in text: 'We have not followed all the stages of degeneration and regeneration in the central end with the same care as in the peripheral end; but the stages we have examined have convinced us that the processes are practically identical in the two ends. The myeline and axis disintegrate and are absorbed for a certain distance; an embryonic fibre is formed from the new protoplasm arising from the nuclei, and in this a myeline sheath is first formed into which an axis cylinder penetrates as an outgrowth from the end of the old axis. Various examnples of this formation of a new fibre within the sheath of the old are shown in Figs. 62 to 72. In many cases, in the central end, when union was not made or when difficult union was made as in cross sutures, an old fibre was found to terminate in a bunch of two or more new fibres (see Figs. 62 and 63), usually coiled round one another so that they could not be disentangled.' (382)
Figs. 63, 65 and 67 in text: 'Still another interesting fact is shown by the teased preparations of the central end of the same nerve, and that is, that the degenerative changes in the central end, when union is not made, apparently keep on progressing centripetally at a slow rate. Figs. 63, 65 and 67 give good illustrations of this fact.' (383)
Figs. 63-65 and 69 in text: 'Cross sections of the central stump of the human ulnar nerve operated upon by Dr Mc Graw for secondary suture, six and a-half months after injury, confirmed the results which were obtained by teasing (see Fig. 69). The section was rnade through the bulbous enlargement of the central stump. At the level of the section no normal medullated fibres were found, though occasionally a cross section of a smaller fibre with some remnant of the axis was seen. In other places a bundle of small fibres was found of the same area as one of these enlarged fibres, and at still other spots intermediate stages were seen showing an enlarged fibre surrounded by small new fibres in the same sheath. In this case the bulbous enlargement was undoubtedly caused by an increase in the nerve fibres as well as in the epineural connective tissue. If the cross section described is compared with the teased preparations, Figs. 63, 64, 65, made from the central end of the same nerve the way in which an old fibre makes connection with a bundle of new fibres lying in the same sheath will be more readily understood. One can understand from the teased preparations how in the cross section a portion of the myeline of an old fibre may be obtained surrounded by a number of newly-formed fibres in the same sheath. ' (383)
Figs. 64 and 65 in text: 'Similar preparations were obtained from the central end of the ulnar, in a dog, which had been severed 75 days before the examination was made, and had not been allowed to unite with the peripheral end. In some of the specimens from this latter nerve the mode of union of the axis cylinder in the newly regenerated fibre with the axis in the old fibre is clearly shown (Figs. 70, 71, and 72). Fig. 71 is particularly instructive when compared with Figs. 64 and 65. The new axis cylinder is seen to escape the swollen end of the old fibre and to penetrate the inyeline some distance beyond this point in order to reach the old axis. Fig. 72 shows the end of an old axis cylinder enlarged and sending out a new axis.' (383-384) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
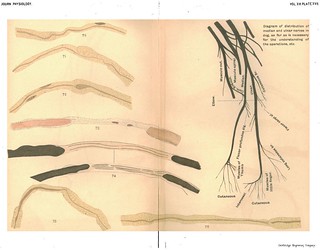
Explanation of Plate XVII (figs. 70-76): 'Figs. 70, 71, 72. To show the connection between the axis cylinder of the old and the new fibre. Compare 71 with figs. 64, 65. Exp. XXII. Central ulnar after 75 days, no union. Picric acid and haematoxylin. Fig. 73. To show the formation of two embryonic fibres in a fibre of central end near the wound, connecting a distal piece which has not degenerated. Central end of human ulnar 61 months after injury. Gold and haematoxylin. Fig. 74. To show intercalated pieces of newly formed fibres without degeneration of distal end. Exp. XVII. L. S. 4 weeks. Wound. Osmic acid and haematoxylin. Fig. 75. The same, from central end of ulnar (Exp. XXII.) 75 days after section, shows the axis cylinder in the intercalated piece and its connection on each side with the intact axis of old fibre. Picric acid and haematoxylin. Fig. 76. The same. Exp. XXII. Central end of ulnar after 75 days. Picric acid and haematoxylin.' (406)
Figs. 70-72 in text: 'We have not followed all the stages of degeneration and regeneration in the central end with the same care as in the peripheral end; but the stages we have examined have convinced us that the processes are practically identical in the two ends. The myeline and axis disintegrate and are absorbed for a certain distance; an embryonic fibre is formed from the new protoplasm arising from the nuclei, and in this a myeline sheath is first formed into which an axis cylinder penetrates as an outgrowth from the end of the old axis. Various examnples of this formation of a new fibre within the sheath of the old are shown in Figs. 62 to 72. In many cases, in the central end, when union was not made or when difficult union was made as in cross sutures, an old fibre was found to terminate in a bunch of two or more new fibres (see Figs. 62 and 63), usually coiled round one another so that they could not be disentangled.' (382) 'Similar preparations were obtained from the central end of the ulnar, in a dog, which had been severed 75 days before the examination was made, and had not been allowed to unite with the peripheral end. In some of the specimens from this latter nerve the mode of union of the axis cylinder in the newly regenerated fibre with the axis in the old fibre is clearly shown (Figs. 70, 71, and 72). Fig. 71 is particularly instructive when compared with Figs. 64 and 65. The new axis cylinder is seen to escape the swollen end of the old fibre and to penetrate the inyeline some distance beyond this point in order to reach the old axis. Fig. 72 shows the end of an old axis cylinder enlarged and sending out a new axis.' (383-384)
Figs. 73-76 in text: 'In conclusion we wish to speak briefly of an appearance often seen in the central end of the nerve, whether or not union is made with the peripheral end, an appearance which has been frequently described by other observers, but which has not been explained, as far as we are aware. This phenomenon consists in the intercalation of a segment of one or even two new fibres between the ends of an old fibre which has not undergone degeneration. (See Figs. 73, 74, 75, 76.) The noteworthy thing is the apparent exception to the general rule that the distal portion of a severed fibre always degenerates.' (384) |
||||||||
