- Creation
-
Date: 1892
- Current Holder(s)
-

- No links match your filters. Clear Filters
-



-
-

-
 Cited byW.H. Howell and G.C. Huber, 'A Physiological, Histological and Clinical Study of the Degeneration and Regeneration in Peripheral Nerve Fibres', Journal of Physiology 13 (5) (1892), pp. 335-406
Cited byW.H. Howell and G.C. Huber, 'A Physiological, Histological and Clinical Study of the Degeneration and Regeneration in Peripheral Nerve Fibres', Journal of Physiology 13 (5) (1892), pp. 335-406
Tags: haemotoxylin, osmic acid, gold
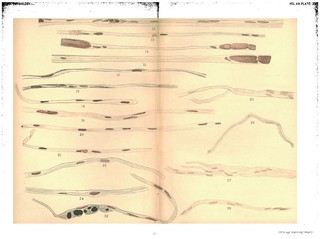
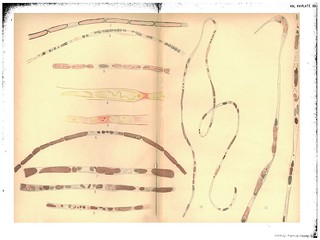
Description:Explanation of Plate XIV (figs. 29-42):
'Fig. 29. Cross-section of human ulnar peripheral end 6 1/2 months after section, union with central end not made, To show cross-section of bundles of embryonic fibres and the appearance of .the nuclei (only one bundle filled in). Gold and haematoxylin.
Fig. 30. The same. Stained in haematoxylin alone.
Fig. 31. Cross-section of pelipheral ulnar 4 weeks after division. After suture but before complete regeneration. To show the resemblance of the nuclei and their nucleoli to cross-sections of young nerve fibres. Exp. XVII. 4 weeks. L. S. Gold and haematoxylin.
Figs. 32, 33. Embryonic fibres in peripheral ulnar of rabbit, 7 weeks after section without union to central end. To show apparent attempt at the formation of myeline. Exp. XXVI. Osmic acid and haematoxylin.
Figs. 34, 35. To show the apparent beginning of a myeline transformation in the protoplasm of the embryonic fibres. Exp. XXVIII. 3 weeks. From branch to flexor profundus. Osmic acid and haematoxylin.
Figs. 36, 37. The first, to show the isolated drops of newly-formed myeline, the second, to show the varicose tube formed by their union. (Portions of the same fibre.) Exp. XXVIII. 3 weeks. Just below wound. Gold and haematoxylin.
Fig. 38. To show the isolated drops of newly-formed myeline and the processes which unite them. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin.
Figs. 39 and 40. To show the same and the staining of the balls of myeline in the haematoxylin. Each of the fibres shows a few fragments of old myeline unabsorbed lying near the nuclei. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin.
Figs. 41 and 41a. The same. To show discontinuous formation of the myeline tube. Exp. XXVIII. Below wound. Osmic acid and haematoxylin.
Fig. 42. The same. Exp. XXVIII. Below wound. Osmic acid and haematoxylin.' (403-404)
Figs. 29-31 in text:
'Our longest experiments upon a nerve cut but not sutured, and in which reunion was prevented by removing a piece of suitable length, were experiment XXVI., upon a rabbit examined 70 days after the operation, and XXII., central ulnar of dog, 75 days. In addition, however, we have had the opportunity of examining a portion of the peripheral end of a human ulnar nerve which had been cut accidentally six and a-half months previously. We owe the opportunity to the kindness of Dr T. A. Mc Graw, who in an operation for secondary suiture removed small portions of the central and peripheral ends and sent them to us in Mueller's solution. Figs. 25, 26, 27, 28 are from drawings of specirmens prepared from the peripheral end of this nerve after treatment with the potash gold stain of Freud and Böhmer's haematoxylin. They show that regeneration had advanced to the stage of the formation of embryonxic fibres, but that there were no signs of an axis cylinder or myeline sheath. Similar preparations were obtained from the animals upon which section without suture was made. Cross sections of the peripheral end of the human nerve are shown in Fig. 29, stained with gold and haematoxylin and in Fig. 30, stained in haematoxylin alone. The drawing in Fig. 29 was made from a portion of the nerve in which some of the nuclei show a central nucleolus in cross section, simulating the appearance of. new nerve fibres with a central axis. This is better shown in Fig. 31, taken from the ulnar nerve of a dog after sutture but before regeneration.' (376-377)
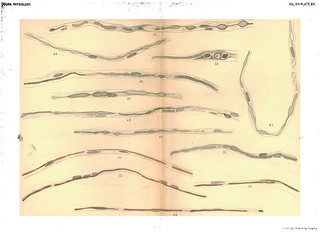
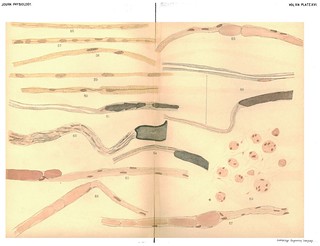
Figs. 35-41 in text:
'Whatever theory of the immediate cause of the formation of the myeline may be the true one, there can be no doubt that it is first found as disconnected drops. These may afterwards become united by slender processes to form a bead-like string which sooner or later elongates to an even tube, or the drops mnay first elongate to form cylindrical segments which eventually unite to form continuous, delicate tubes of myeline. Both of these processes, with the intermediate stages, are shown in Figs. 36 to 55 better than we could describe them in words. What becomes of the numerous nuclei scattered along the embryonic fibre it is not possible to say, other than that they disappear. In some cases, as in Figs. 38, 39, 42, they disappear rapidly as the myeline tube is formed, while in other cases they persist for a much longer time, the newly-formed myeline tube winding in and out among them in a very beautiful manner (see Figs. 37, 51, 55). We can only suppose that they disappear by absorption, as their nutritive relations with reference to the protoplasmic contents of the fibre become less and less important. With reference to the nodes and internodes of Ranvier, it is evident that no simple hypothesis, such as the development of each internode from a single cell, will fit the facts as they appear in regenerating fibres. The developing internodes and nodes are plainly shown in Figs. 51 to 55, but why the ends of the internodal tubes do not fuse together is difficult to explain.' (379-380)
Figs. 36-41 in text:
'After the formation of the embryonic fibres the additional steps necessary for a complete regeneration consist in the production of a new myeline sheath and a new axis cylinder... Figs. 36, 37, 39, 40 and 41 give an idea of how the myeline first appears. As shown in these figures, it appears first as irregular deposits in the protoplasm of the embryonic fibres, and usually first in the immediate neighbourhood of the nuclei. Delicate prolongations of the myeline are often seen running from one small mass of the myeline to another, and eventually these latter become connected together, forming a varicose tube, shown in various stages in Figs. 36 to 51. With reference to the immediate origin of these deposits, we must confess ourselves in doubt. In many cases we obtained specimens such as are shown in Figs. 34 and 35, in which clear bead-like spaces are seen forming in the protoplasm. The general arrangement of these spaces strongly suggests that they mark the beginning of the transformation of the protoplasm to myeline; a step farther and the chemical change will be such as to give the usual staining with osmic acid or gold as seen in Figs. 36-38, etc. On the other hand, the first deposits of myeline, especially when stained in osmic acid, often bear a striking resemblance to the nuclei, both in shape and in the fact that they show a rather distinct staining with the haematoxylin. (See Figs. 38, 39, 40, 42, etc.) There is no doubt that as the myeline forms many of the numerous nuclei scattered along the fibre disappear.' (378-379)
-
-
-
-
-