Tag: gold
Category: All General/Other Stained with
No results
|
|
|
cites |
 |
|
  |
|||
|
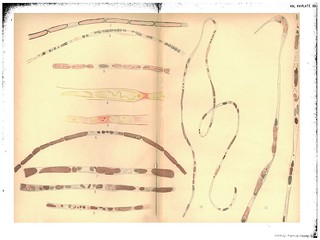
Explanation of Plate XII (figs. 1-11): 'The figures with one exception were drawn under a zeiss D and ocular 4. The outlines were filled in with the Camera Lucida and an attempt made to colour the drawings like the particular specimen under examination. Fig. 1. To show the first segmentation of the myeline in degeneration. Each segment completely enclosed in myeline, with a central piece of the axis cylinder. Experiment XX. 2 days. R. S. middle of forearm. Osmic acid and haematoxylin. Fig. 2. To show the secondary fragmentation of the myeline (extreme case). Exp. XX. 2 days. L S. middle of forearm. Osmic acid and haematoxylin. Fig. 3. To show the remnants of the disintegrating axis in the segments of myeline. Exp. XIII. 4 days. R. S. 3 in. below wound. Gold and haematoxylin. Nucleus stained blue in haematoxylin. Fig. 4. The same. Exp. IIl. 7 days. R. S. Below wound 2 in. Picric acid and fuchsin stain. Fig. 5. The same. Exp. III. 7 days. R. S. Below wound 2 in. Picric acid and fuchsin stain. Fig. 6. To show the more rapid fragmentation of the myeline in the neighbourhood of the nuclei. Exp. III. 7 days. Gold and haematoxylin. Nucleus stained blue. Fig. 7. To show the multiplication and migration of the nuclei of the sheath and greater absorption near nuclei. Exp. III. 7 days. Gold anld haematoxylin. Fig. 8. The same. Exp. III. 7 days. Gold and haematoxylin. Fig. 9. An instance of apparent indirect division of nucleus. The progressing fragmentation of myeline. Exp. III. 7 days. Gold and haematoxylin. Fig. 10. (Drawn under obj. C. oc. 4). To show fragmentation of myeline especially at nuclei, and the material containing fragments of myeline which fills the fibre. Exp. III. 7 days. Gold and haematoxylin. Fig. 11. To show progressive absorptionl of myeline; the proliferation of the nuclei; the position of the nuclei with reference to the balls of myeline; and the collapse of the empty sheath. Exp. XI. 9 days. R. S. middle of forearm. Gold and haematoxylin.' (402)
Fig. 1 in text: 'The segments first formed in degeneration, unlike the normal appearance of the segments of Lantermann seen in fibres after the action of osmic acid, have a complete envelope of the myeline, as shown in Fig. 1, and the small interspaces between them are filled with a colourless material. In each of these segments is a piece of the original axis cylinder, at this time apparently unchanged. Many of the older histologists have asserted that the axis remains unaffected during the degeneration. On the contrary, even at this early stage the breaking of the myeline into segments is accompanied by, or causes a simultaneous breaking of the axis.' (371)
Figs. 2-8 in text: 'Shortly after the first cylindrical segments of myeline and axis are formed an irreaular fragmentation occurs in these segments in most parts of the fibre. The segmetnts break up into smaller or larger irregular pieces or balls, an extreme case of which is shown in Fig. 2. Very frequently in these irregular masses of myeline remnants of the axis cylinder can be clearly distinguished in all stages of disintegration as shown in Figs. 3, 4, 5. It should be stated that some of the large segments frequently persist in different portions of a fibre long after tlle other segments have broken into small pieces and become partially absorbed,-the process is very irregular. It is at the time that this breaking up of the cylindrical segments into smaller fragments becomes apparent that the increase in the size of the nuclei of the sheath and the growth of the protoplasm surrounding these rnuclei become clearly marked. In fact this secondary fragmentation is always visible first in the neighbourhood of the nuclei as shown in Figs. 6, 7, 8. The large rounded nuclei lie in the middle of the fibre, and close to them are the small drops or balls of myeline. For this reason we believe that one cause of this secondary fragmentation is to be found in the absorption which takes place under the influence of the nucleus and its suirrounding protoplasm.' (372)
Fig. 6 in text: 'As evidence of the actual absorption which is beginning to take place one finds, at this time, say the seventh day, in the fibres stained with osmic acid or gold, some of the balls of myeline in the neighbourhood of the nuclei left colourless by the stain, see Fig. 6. In the later stages the same fact may frequently be noticed at different parts of the fibre and always most clearly near the nuclei. See Fig. 11.' (372-373)
Figs. 7-8 in text: 'An interesting fact in connection with the inultiplication of the nuclei is the way in which they migrate. In the beginning, of course, there is a single nucleus to each internode. At the time the secondary fragmentation of the myeline is fully under way one frequently finds a number of nuclei in the space which an internode would occupy. Sometimes they are in pairs as though from a recent division, but in other cases one or more large masses of myeline will be found between. See Figs. 7 and 8. This latter appearance has been used to support the theory that there are several nuclei present in the internode in a normal fibre, but that they are hidden by the myeline. Such a view as this it is not necessary to consider at present. The only explanation of the appearance described that seems reasonable is that after division the nuclei migrate or may migrate to some distance and start the process of absorption at a new place.' (373)
Fig. 9 in text: 'By the 7th day a very active proliferation of the nuclei of the sheath has begun. The increase in their number is very striking. Our methods of hardening were not such as to show the method of multiplication of the nuclei, though we often found nuclei showing a dumb-bell form, i.e., an elongated nucleus constricted in the middle as though multiplying by direct division. There can be little doubt that the division is indirect, as we sometimes found, even after hardening in Mueller's solution, nuclei like that pictured in Fig. 9 which evidently represents a badly preserved mitosis.' (
Figs. 10-11 in text: 'From the 7th day to the 14th day the process of absorption of the balls of myeline with their contained fragments of axis cylinder goes on actively, yet quite irregularly. Fig. 10 from a nerve after 7 days shows very well the breaking-up of the myeline and the formation in between, especially at the nuclei, of an apparently liquid substance in which are contained numerous fragments of the old myeline. As the absorption progresses and the fragments of myeline become smaller and less numerous the direct participation of the nuclei in the process becomes more evident. The nuclei are much more numerous and are found clustered in and about the remaining balls of myeline as shown in Figs. 11 and 12, representing a degeneration of 9 and 14 days respectively. One often sees bits of the myeline partially imbedded in a nucleus, and this appearance is found from this time on well into the later stages of regeneration, as long, in fact, as any of the myeline remiains unabsorbed. After 14 days absorption has gone so far that long stretches may be found, as shown in Figs. 12 and 13, in which only small fragments of myeline are present. At such places the fibre consists of a homogeneous, apparently liquid substance lying in the old sheath, and of many nuclei, often in pairs or groups, the latter giving indication of an active proliferation. Yet at this time, 14 days, and even later, one sees many fibres in which the absorption has lagged behind the condition of what may be considered a typical fibre of this period. In one and the same fibre places will be found in which absorption has made rapid progress in spots, all the myeline having disappeared, while in other spots the large cylindrical segments have suffered scarcely any change. Examples of this are pictured in Figs. 14 and 15. However irregularly the process may go on the final outcome is the complete absorption of the remnanits of the old myeline and axis; though as we have said before, balls of the myeline may be found in certain fibres long after this period, even at the time when fully formed new fibres have beeu produced. As the absorption proceeds the old sheath collapses more or less. It seems at first to contain a liquid material with some debris of the old myeline, but this too finally disappears and the beginning of the actual process of regeneration is inaugurated by the formation of new protoplasmic material around the numerous nuclei contained in the fibres.' (373-374) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
Explanation of Plate XIV (figs. 29-42): 'Fig. 29. Cross-section of human ulnar peripheral end 6 1/2 months after section, union with central end not made, To show cross-section of bundles of embryonic fibres and the appearance of .the nuclei (only one bundle filled in). Gold and haematoxylin. Fig. 30. The same. Stained in haematoxylin alone. Fig. 31. Cross-section of pelipheral ulnar 4 weeks after division. After suture but before complete regeneration. To show the resemblance of the nuclei and their nucleoli to cross-sections of young nerve fibres. Exp. XVII. 4 weeks. L. S. Gold and haematoxylin. Figs. 32, 33. Embryonic fibres in peripheral ulnar of rabbit, 7 weeks after section without union to central end. To show apparent attempt at the formation of myeline. Exp. XXVI. Osmic acid and haematoxylin. Figs. 34, 35. To show the apparent beginning of a myeline transformation in the protoplasm of the embryonic fibres. Exp. XXVIII. 3 weeks. From branch to flexor profundus. Osmic acid and haematoxylin. Figs. 36, 37. The first, to show the isolated drops of newly-formed myeline, the second, to show the varicose tube formed by their union. (Portions of the same fibre.) Exp. XXVIII. 3 weeks. Just below wound. Gold and haematoxylin. Fig. 38. To show the isolated drops of newly-formed myeline and the processes which unite them. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin. Figs. 39 and 40. To show the same and the staining of the balls of myeline in the haematoxylin. Each of the fibres shows a few fragments of old myeline unabsorbed lying near the nuclei. Exp. XXVIII. 3 weeks. Just below wound. Osmic acid and haematoxylin. Figs. 41 and 41a. The same. To show discontinuous formation of the myeline tube. Exp. XXVIII. Below wound. Osmic acid and haematoxylin. Fig. 42. The same. Exp. XXVIII. Below wound. Osmic acid and haematoxylin.' (403-404)
Figs. 29-31 in text: 'Our longest experiments upon a nerve cut but not sutured, and in which reunion was prevented by removing a piece of suitable length, were experiment XXVI., upon a rabbit examined 70 days after the operation, and XXII., central ulnar of dog, 75 days. In addition, however, we have had the opportunity of examining a portion of the peripheral end of a human ulnar nerve which had been cut accidentally six and a-half months previously. We owe the opportunity to the kindness of Dr T. A. Mc Graw, who in an operation for secondary suiture removed small portions of the central and peripheral ends and sent them to us in Mueller's solution. Figs. 25, 26, 27, 28 are from drawings of specirmens prepared from the peripheral end of this nerve after treatment with the potash gold stain of Freud and Böhmer's haematoxylin. They show that regeneration had advanced to the stage of the formation of embryonxic fibres, but that there were no signs of an axis cylinder or myeline sheath. Similar preparations were obtained from the animals upon which section without suture was made. Cross sections of the peripheral end of the human nerve are shown in Fig. 29, stained with gold and haematoxylin and in Fig. 30, stained in haematoxylin alone. The drawing in Fig. 29 was made from a portion of the nerve in which some of the nuclei show a central nucleolus in cross section, simulating the appearance of. new nerve fibres with a central axis. This is better shown in Fig. 31, taken from the ulnar nerve of a dog after sutture but before regeneration.' (376-377)
Figs. 35-41 in text: 'Whatever theory of the immediate cause of the formation of the myeline may be the true one, there can be no doubt that it is first found as disconnected drops. These may afterwards become united by slender processes to form a bead-like string which sooner or later elongates to an even tube, or the drops mnay first elongate to form cylindrical segments which eventually unite to form continuous, delicate tubes of myeline. Both of these processes, with the intermediate stages, are shown in Figs. 36 to 55 better than we could describe them in words. What becomes of the numerous nuclei scattered along the embryonic fibre it is not possible to say, other than that they disappear. In some cases, as in Figs. 38, 39, 42, they disappear rapidly as the myeline tube is formed, while in other cases they persist for a much longer time, the newly-formed myeline tube winding in and out among them in a very beautiful manner (see Figs. 37, 51, 55). We can only suppose that they disappear by absorption, as their nutritive relations with reference to the protoplasmic contents of the fibre become less and less important. With reference to the nodes and internodes of Ranvier, it is evident that no simple hypothesis, such as the development of each internode from a single cell, will fit the facts as they appear in regenerating fibres. The developing internodes and nodes are plainly shown in Figs. 51 to 55, but why the ends of the internodal tubes do not fuse together is difficult to explain.' (379-380)
Figs. 36-41 in text: 'After the formation of the embryonic fibres the additional steps necessary for a complete regeneration consist in the production of a new myeline sheath and a new axis cylinder... Figs. 36, 37, 39, 40 and 41 give an idea of how the myeline first appears. As shown in these figures, it appears first as irregular deposits in the protoplasm of the embryonic fibres, and usually first in the immediate neighbourhood of the nuclei. Delicate prolongations of the myeline are often seen running from one small mass of the myeline to another, and eventually these latter become connected together, forming a varicose tube, shown in various stages in Figs. 36 to 51. With reference to the immediate origin of these deposits, we must confess ourselves in doubt. In many cases we obtained specimens such as are shown in Figs. 34 and 35, in which clear bead-like spaces are seen forming in the protoplasm. The general arrangement of these spaces strongly suggests that they mark the beginning of the transformation of the protoplasm to myeline; a step farther and the chemical change will be such as to give the usual staining with osmic acid or gold as seen in Figs. 36-38, etc. On the other hand, the first deposits of myeline, especially when stained in osmic acid, often bear a striking resemblance to the nuclei, both in shape and in the fact that they show a rather distinct staining with the haematoxylin. (See Figs. 38, 39, 40, 42, etc.) There is no doubt that as the myeline forms many of the numerous nuclei scattered along the fibre disappear.' (378-379) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
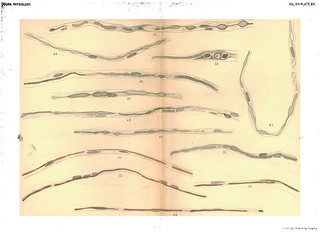
Explanation of Plate XV (figs. 42-55): 'Fig, 43. To show varicose appearance of newly-formed tube of myeline. Exp, XXVIII. 3/4 in. below wound. Osmic acid and haematoxylin. Fig. 44. The same. Exp. XXII. R. S. near wrist. Osmic acid and haematoxylin. Fig. 46. To show the formation of the myeline tube without varicosities. Exp. XXVIII. 3/4 in. below wound. Osmnic acid and haematoxylin. Fig. 47. To show union of isolated drops of myeline to form a tube. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 48. To show discontinuous formation of myeline sheath. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 49, The same. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 50. To show the persistence of balls of old myeline as yet unabsorbed and their position with reference to the nuclei. Exp. XXVIII. Osmic acid and haematoxylin. Nuclei stained in haematoxylin. Fig. 51. To show the formation of a node of Ranvier. Exp. XXVIII. Osmic acid and haematoxylin. Fig. 52. To show the newly-formed myeline tube lying in the embryonic fibre. Exp. IX. 3 weeks. From dorsal cutaneous branch. Gold and haematoxylin. Fig. 53. The same. Exp. IX. L. S. Dorsal cutaneous branch. Gold and haematoxylin. Fig. 54. To show the formation of nodes of Ranvier. Exp. IX. L. S. Dorsal cutaneous branch. Gold and haematoxylin. Fig. 55. To sbow newly-formed myeline tube with node of Ranvier and persistent nuclei. Exp. IX. L. S. 3 weeks. At wound. Gold and haematoxylin.' (404-405)
Figs. 42-55 in text: 'Whatever theory of the immediate cause of the formation of the myeline may be the true one, there can be no doubt that it is first found as disconnected drops. These may afterwards become united by slender processes to form a bead-like string which sooner or later elongates to an even tube, or the drops mnay first elongate to form cylindrical segments which eventually unite to form continuous, delicate tubes of myeline. Both of these processes, with the intermediate stages, are shown in Figs. 36 to 55 better than we could describe them in words. What becomes of the numerous nuclei scattered along the embryonic fibre it is not possible to say, other than that they disappear. In some cases, as in Figs. 38, 39, 42, they disappear rapidly as the myeline tube is formed, while in other cases they persist for a much longer time, the newly-formed myeline tube winding in and out among them in a very beautiful manner (see Figs. 37, 51, 55). We can only suppose that they disappear by absorption, as their nutritive relations with reference to the protoplasmic contents of the fibre become less and less important. With reference to the nodes and internodes of Ranvier, it is evident that no simple hypothesis, such as the development of each internode from a single cell, will fit the facts as they appear in regenerating fibres. The developing internodes and nodes are plainly shown in Figs. 51 to 55, but why the ends of the internodal tubes do not fuse together is difficult to explain.' (379-380)
Fig. 42 in text: 'the first deposits of myeline, especially when stained in osmic acid, often bear a striking resemblance to the nuclei, both in shape and in the fact that they show a rather distinct staining with the haematoxylin. (See Figs. 38, 39, 40, 42, etc.) There is no doubt that as the myeline forms many of the numerous nuclei scattered along the fibre disappear.' (379) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
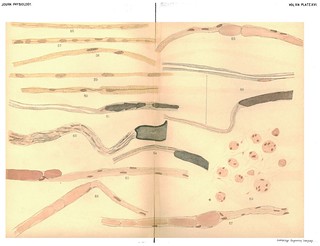
Explanation of Plate XVI (figs. 56-69): Figs. 56, 57, 58, 59. From nerve stained in picric acid atid haematoxylin. Exp. XIX. 7 weeks. R. S. J in. below wound. 59. An embryonic fibre. 58. Newly-formed fibres with myeline and axis. 57 and 56. Newly forming fibres, myeline as a varicose tube. Axis also present but not stained in the interior of the swellings of myeline. Shows how quickly after the formation of the myeline the axis grows down from above. Fig. 60. To show the branching of the axis cylinder where an old fibre passes into several new fibres, central end of ulnar. Exp. XXlI. Cut for 75 days and union with peripheral end prevented. Picric acid and haematoxylin. Fig. 61. To show what seems to be the outgrowth of an axis cylinder from an old fibre toward a new one. New myeline lies in the embryonic fibre as a continuous delicate sheath. Exp. XVII. 4 weeks. R. S. Gold and haematoxyiin. Fig. 62. To show the formation of several new fibres in the degenerated portion of a single old fibre at the central end of wound. Exp. XVII. 4 weeks. L. S. wound of median ulnar suture. Osmic acid and haematoxylin. Fig. 63. The sanme. Human ulnar nerve. Central end of nerve 6 1/2 months after injury. No union with peripheral end. Gold and haematoxylin. Fig. 64. To show junction of old with new fibre at the central end. Exp. XVIII. R. S. wound. Osmic acid and haematoxylin. Fig. 65. The same. Human ulnar. Central end 6 1/2 months after section and no union with peripheral end. The continuation of the protoplasm of the "embryonic fibre" into the hypertrophied protoplasm surrounding the nuclei of the old fibre, and the fragmentation of the end of the old myeline to be specially noted. Gold and haematoxylin. Fig. 66. To show junction of old and new fibres, and the new fibre with its thin myeline sheath lying in the protoplasm of the "embryonic fibre." Exp. XVII. R. S. wound. Gold and haematoxylin. Fig. 67. To show junction of old and new fibres. Central end of human ulnar, 6 1/2 months after injury and no union with peripheral end. The fragmentation of the myeline and the hypertrophy of the protoplasm round the nuclei of the old fibre to be noted. Gold and haematoxylin. Fig. 68. The same. Fig. 69. Cross-section through the bulbous enlargement of central end of human ulnar, 6 1/2 months after injury and no union with central end. Shows the increase in the endoneural connective tissue as well as in the nerve fibres. The fibre marked + shows cross section of new fibres in same sheath with old. Compare with figs. 63, 65.' (405-406)
Figs. 56-57 in text: 'when the new myeline is evidently just formed no definite axis can be demonstrated by the gold stain. With the picric acid and haematoxylin stain, on the contrary, the newly formed axis is clearly seen, even at that early stage in the formation of the myeline tube when it exists as a string of bead-like swellings (see Figs. 56 and 57). It follows then that though the myeline sheath probably begins to form before the axis cylinder can be distinguished, the latter appears shortly afterward, before the new fibre has gone far in its development.' (380-381)
Fig. 61 in text: 'Very many apparent examples of this branching of the axis were found in this specimen, but owing, to the intricate way in which the fibres were twisted and the possibilities of deception arising therefrom, the connection of the old and new axes could not be satisfactorily followed. The example given in the drawing, and some others, were, however, quite distinct and seem to us to give fairly satisfactory histological proof that in regeneration the new axis cylinders are outgrowths from the axes of the uninjured fibres of the cenitral end. Fig. 61 gives an apparent example from another experiment of the outgrowth of the axis.' (382)
Figs. 62-69 in text: 'We have not followed all the stages of degeneration and regeneration in the central end with the same care as in the peripheral end; but the stages we have examined have convinced us that the processes are practically identical in the two ends. The myeline and axis disintegrate and are absorbed for a certain distance; an embryonic fibre is formed from the new protoplasm arising from the nuclei, and in this a myeline sheath is first formed into which an axis cylinder penetrates as an outgrowth from the end of the old axis. Various examnples of this formation of a new fibre within the sheath of the old are shown in Figs. 62 to 72. In many cases, in the central end, when union was not made or when difficult union was made as in cross sutures, an old fibre was found to terminate in a bunch of two or more new fibres (see Figs. 62 and 63), usually coiled round one another so that they could not be disentangled.' (382)
Figs. 63, 65 and 67 in text: 'Still another interesting fact is shown by the teased preparations of the central end of the same nerve, and that is, that the degenerative changes in the central end, when union is not made, apparently keep on progressing centripetally at a slow rate. Figs. 63, 65 and 67 give good illustrations of this fact.' (383)
Figs. 63-65 and 69 in text: 'Cross sections of the central stump of the human ulnar nerve operated upon by Dr Mc Graw for secondary suture, six and a-half months after injury, confirmed the results which were obtained by teasing (see Fig. 69). The section was rnade through the bulbous enlargement of the central stump. At the level of the section no normal medullated fibres were found, though occasionally a cross section of a smaller fibre with some remnant of the axis was seen. In other places a bundle of small fibres was found of the same area as one of these enlarged fibres, and at still other spots intermediate stages were seen showing an enlarged fibre surrounded by small new fibres in the same sheath. In this case the bulbous enlargement was undoubtedly caused by an increase in the nerve fibres as well as in the epineural connective tissue. If the cross section described is compared with the teased preparations, Figs. 63, 64, 65, made from the central end of the same nerve the way in which an old fibre makes connection with a bundle of new fibres lying in the same sheath will be more readily understood. One can understand from the teased preparations how in the cross section a portion of the myeline of an old fibre may be obtained surrounded by a number of newly-formed fibres in the same sheath. ' (383)
Figs. 64 and 65 in text: 'Similar preparations were obtained from the central end of the ulnar, in a dog, which had been severed 75 days before the examination was made, and had not been allowed to unite with the peripheral end. In some of the specimens from this latter nerve the mode of union of the axis cylinder in the newly regenerated fibre with the axis in the old fibre is clearly shown (Figs. 70, 71, and 72). Fig. 71 is particularly instructive when compared with Figs. 64 and 65. The new axis cylinder is seen to escape the swollen end of the old fibre and to penetrate the inyeline some distance beyond this point in order to reach the old axis. Fig. 72 shows the end of an old axis cylinder enlarged and sending out a new axis.' (383-384) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
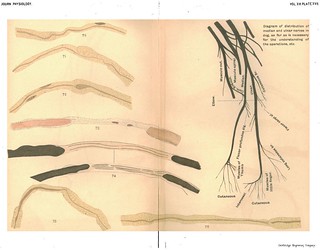
Explanation of Plate XVII (figs. 70-76): 'Figs. 70, 71, 72. To show the connection between the axis cylinder of the old and the new fibre. Compare 71 with figs. 64, 65. Exp. XXII. Central ulnar after 75 days, no union. Picric acid and haematoxylin. Fig. 73. To show the formation of two embryonic fibres in a fibre of central end near the wound, connecting a distal piece which has not degenerated. Central end of human ulnar 61 months after injury. Gold and haematoxylin. Fig. 74. To show intercalated pieces of newly formed fibres without degeneration of distal end. Exp. XVII. L. S. 4 weeks. Wound. Osmic acid and haematoxylin. Fig. 75. The same, from central end of ulnar (Exp. XXII.) 75 days after section, shows the axis cylinder in the intercalated piece and its connection on each side with the intact axis of old fibre. Picric acid and haematoxylin. Fig. 76. The same. Exp. XXII. Central end of ulnar after 75 days. Picric acid and haematoxylin.' (406)
Figs. 70-72 in text: 'We have not followed all the stages of degeneration and regeneration in the central end with the same care as in the peripheral end; but the stages we have examined have convinced us that the processes are practically identical in the two ends. The myeline and axis disintegrate and are absorbed for a certain distance; an embryonic fibre is formed from the new protoplasm arising from the nuclei, and in this a myeline sheath is first formed into which an axis cylinder penetrates as an outgrowth from the end of the old axis. Various examnples of this formation of a new fibre within the sheath of the old are shown in Figs. 62 to 72. In many cases, in the central end, when union was not made or when difficult union was made as in cross sutures, an old fibre was found to terminate in a bunch of two or more new fibres (see Figs. 62 and 63), usually coiled round one another so that they could not be disentangled.' (382) 'Similar preparations were obtained from the central end of the ulnar, in a dog, which had been severed 75 days before the examination was made, and had not been allowed to unite with the peripheral end. In some of the specimens from this latter nerve the mode of union of the axis cylinder in the newly regenerated fibre with the axis in the old fibre is clearly shown (Figs. 70, 71, and 72). Fig. 71 is particularly instructive when compared with Figs. 64 and 65. The new axis cylinder is seen to escape the swollen end of the old fibre and to penetrate the inyeline some distance beyond this point in order to reach the old axis. Fig. 72 shows the end of an old axis cylinder enlarged and sending out a new axis.' (383-384)
Figs. 73-76 in text: 'In conclusion we wish to speak briefly of an appearance often seen in the central end of the nerve, whether or not union is made with the peripheral end, an appearance which has been frequently described by other observers, but which has not been explained, as far as we are aware. This phenomenon consists in the intercalation of a segment of one or even two new fibres between the ends of an old fibre which has not undergone degeneration. (See Figs. 73, 74, 75, 76.) The noteworthy thing is the apparent exception to the general rule that the distal portion of a severed fibre always degenerates.' (384) |
||||||||
