- External URL
- Creation
-
Creator (Definite): George Lovell GullandDate: 1896
- Current Holder(s)
-

- No links match your filters. Clear Filters
-



-
 Cites
Cites Plate V, Journal of Physiology 19 (5-6) (1896). Figs. 1-16 from G.L. Gulland, 'On the Granular Leucocytes'.
Plate V, Journal of Physiology 19 (5-6) (1896). Figs. 1-16 from G.L. Gulland, 'On the Granular Leucocytes'.
Description:Explanation of Plate V (figs. 1-16):
'The objective used in all cases except Fig. 41 was Zeiss's apochromatic homogeneous immersion 2.0 mm. equivalent focus and 1.40 mm. aperture, and the eyepiece was 18 compensating, in all cases but Figs. 3, 6 and 41. Figs. 3 and 6 were drawn with eyepiece 8 compensating. Fig. 41 with eyepiece 2 compensating and Zeiss objective DD.
The magnification for the majority of the figures is x 2250; for Figs. 3 and 6 x 1,000, for Fig. 41, about x 200.
Fig. 1. Hyaline leucocyte from blood of newt, moving slightly, astrosphere very evident. Iron-haematoxylin.
Fig. 2. Small oxyphile leucocyte from newt's blood, mitoma very evident, nucleus rather over-stained. Iron-haematoxylin.
Fig. 3. Eosinophile cell from newt's blood with spherical pseudopod. n = nucleus, which was over-stained. Iron-haematoxylin, x 1,000.
Fig. 4. Basophile cell, very small, from rabbit's mesentery. n = nucleus, c = centrosomes nearly equal in size, the centrodesmosis took up the stain only very faintly. Methylene-blue.
Fig. 5. Detached pseudopod from eosinophile cell of newt's blood; the optical section of the middle is drawn; the points unattached represent the places where the mitoma passed out of view, either upwards or downwards. Methylene-blue, Bordeaux.
Fig. 6. Eosinophile cell of newt's blood with spherical pseudopod attached by thin thread. n = nucleus over-stained. Bordeaux, iron-haematoxylin.
Fig. 7. Hyaline leucocyte of frog's blood. as = astrosphere with two centrosomes very unequal in size; some of the microsomes in the ring bounding the astrosphere have retained the stain; n = nucleus over-stained. The cytomitoma though visible was not evident enough to be drawn in detail. Bordeaux, iron-hoematoxylin.
Fig. 8. Oxyphile leucocyte (amphophile) from bone-marrow of rabbit; mitoma very evident, microsomes small. Iron-haematoxylin.
Fig. 9. Hyaline leucocyte from bone-marrow of rabbit; microcentre with four centrosomes, mitoma fairly evident. Iron-haematoxylin.
Fig. 10. Eosinophile cell from bone-marrow of rabbit; astrosphere well seen, two small centrosomes. Iron-haematoxylin.
Fig. 11. Transition form between oxyphile and eosinophile cells from bone-marrow of rabbit, evidently very small part of the cell. Iron-haematoxylin.
Fig. 12. Eosinophile cell from bone-marrow of rabbit; astrosphere well seen. The section has evidently passed right through the middle of the cell, and through the middle of a spectacle-shaped nucleus. Iron-haematoxylin.
Fig. 13. Part of an eosinophile cell from bone-marrow of rabbit; the fainter microsomes were those slightly out of focus, above or below, when drawing the nucleus. Iron-hsematoxylin.
Fig. 14. Eosinophile cell from tongue of frog; fainter microsomes slightly out of focus. Iron-haematoxylin.
Fig. 15. Part of an eosinophile cell from tongue of a frog; no nucleus in the section, mitoma specially well seen. Iron-heematoxylin.
Fig. 16. Eosinophile cell from hen's ovary; astrosphere well seen; radial (?) arrangement of microsomes. Section probably cut as in Fig. 12. Iron-heematoxylin.' (415-416)
Figs. 1, 7 and 9 in text:
'It must not... be thought that the hyaline cell-body is structureless. The constant presence of the microcentre alone would prove the contrary, and though no hint of structure can be seen in the lymphocytes, or even in the smaller hyaline cells, yet as they become larger, one can see the ordinary network of protoplasm (see Figs. 7 and 9). In the hyaline cells the nucleus may remain round or may alter its shape according to Heidenhain's law, that is, according to the relation of its diameter to the diameter -of the whole cell (Figs. 1, 7, 9). The process of change from the small hyaline cell to the finely granular oxyphile cell is associated with an increase in the visibility of the mitoma. It is this, in the early stages, rather than the actual appearance of granules that gives the finely granular appearance to the cell-body. Along with this goes the deformation of the nucleus by amoeboid movements, and the ordinary polymorphous form is soon reached. When these forms are seen in the blood the cells are generally approximately spherical, and the microcentre is situated in the middle of the cell (Figs. 18 and 19). The centrosomes are small, too small for one to be sure of their number as a rule, and I have found them stain better with the unmodified Ehrlich-Biondi stain than with iron-haematoxylin. The limits of the astrosphere are not usually well-marked, except sometimes in the larger cells (Fig. 19).' (399)
Figs. 2 and 8 in text:
'Minute as these microsomes are it can already be made out that they vary in size to a certain extent; on the whole, the largest microsomes are towards the periphery of the cell. In the larger cells of this stage the interstitial substance between the threads of the mitoma becomes increased in amount and the strands are consequently placed further apart. The difference is seen in the contrast between Figs. 18 and 19 and in Figs. 2 and 8.' (399)
Figs. 2, 8 and 11 in text:
'The transition-forms between the finely granular and the coarsely granular acidophile cells are seen much more frequently in the bone-marrow than in the blood, and it seems certain that both from this source and from mitotic division the main source of the eosinophile cells is in the bone-marrow. They must arise elsewhere however in abundance, for both Schaffer [ref: '(7)' ['Schaffer, J. Centrlbl. f. d. med. Wiss. 1891.']] and I [note: '(20) ['Gulland, G. L. Journ. of Path. and Bacter. 1894. ']] have shown that they are present in the thymus and in lymphatic glands before either bone or bone-marrow is properly formed at all, and Engel [note: '(19)' ['Engel, C. S. Arch. f. mikr. Anat. LXIV. 1894.']] has seen them in the chick's blood on the 5th day of incubation. In the transition-forms (see Figs. 2, 8, 11) there is little in the general shape of the cell and nucleus to distinguish them from- the preceding stage. The real difference is the fact that the mitoma has become much more visible and still more widely spaced, and th-at some of the microsomes are much larger than others. These larger microsomes take up eosin and iron-haematoxylin much more deeply than the smaller granules, and it is at this stage too that the threads of the mitoma can first be said to stain satisfactorily with iron-hoematoxylin (see Fig. 11).' (399-400)
Figs. 3, 5-6 in text:
'The mitoma in the eosinophile cell is especially well seen in those of the newt's blood. These cells are markedly amceboid and have the habit of throwing out circular pseudopodia which are often counected to the main part of the cell only by a very delicate thread. Two such cells are shown in Figs. 3 and 6, and it is evident that the threads are often broken through and the spherical portion of the cell-body set free, as the blood contains a large number of them. The mitoma in these is often very clear, as may be seen from Fig. 5 and from Fig. 17, where one of these bodies has been ingested by a "hyaline" leucocyte.' (401)
Fig. 4 in text:
'I shall best be able to describe the changes through which they pass by tracing the cells from the original lymphocyte or hyaline transitionform, from which they are derived. The first change consists not so much in an enlargement of the microsomes as in the superior visibility of the mitoma. As in the eosinophile cells the cell-body becomes larger, and as there the mitoma takes up acid stains, so here it takes up basic stains (Fig. 4). As the cell increases in size, the mitoma retains its visibility and the microsomes become larger and assume the ordinary appearance of granules. These are at first few and scattered irregularly through the cell: some microsomes have increased in size, others have not; the mitoma is always most visible near the microsomes which have taken up the stain. At this stage, illustrated by Figs. 21-23, from the blood, and Figs. 24-27, from the mesentery, the granules always differ markedly in size, much more so than at later stages. They are always uniform in staining, and curiously enough, the very small ones do not often show the metachromatic reaction with methylene-blue. The cells in the blood are generally spherical, but those in connective tissue soon show an irregularity in shape suggestive of amoeboid movements; they put forth those long flat processes which are so characteristic of the larger cells. In Figs. 26 and 27, for instance, all the granules to be seen in the cell are shown, and they were all lying almost in one plane, so that hardly any focussing was required to see them. They must therefore have been lying in a very thin layer of protoplasm, and from the constancy of this appearance we may infer that the spaces through which these leucocytes have to pass between the planes of connective tissue in the mesentery must be very shallow. The rounder cells are generally found in places where, as near the larger blood vessels, the mesentery becomes thicker and allows more space; the elongated cells are generally in the thin parts of the structure.' (406)
'Westphal [ref: '(4)' ['Ehrlich, P. Farbenanalytische Untersuchungen &c. Gesammelte Mittheilungen. Pt. I. 1891.']] distinguished three main forms of the large basophile cells, flat, spherical, and spindle-shaped, with two or more processes; he remarks however that this is purely a classification for convenience, as transition-forms of every kind are to be seen also. This I can fully confirm, as I can also his dicta that "in individuals of the same species and same age the distribution, number and size of the cells are similar in similar organs," and that in individuals of different species there are great differences in the number &c., of the cells in the same organs. I may add that certain species show a tendency to have cells of one or other shape or size. The mesentery of the rabbit, for instance, contains, speaking generally, small cells, mostly spherical (Figs. 4, 24, 25); of the guinea-pig, larger spindle-shaped cells (Figs. 29 and 30); of the rat, enormous round and flat cells (Figs. 31, 32); while that of the newt has huge spindle cells which send off often several secondary branches from the primary ones (Fig. 41).' (403-404)
Fig. 5 in text:
'As there is neither nucleus nor centrosome in these portions they soon degenerate, but like eosinophile cells elsewhere they retain their integrity to the last. Fig. 5 shows one of them apparently soon after its detachment from the leucocyte, while the mitoma is still very evident. Indeed, it has seemed to me as although in the earlier stages of degeneration the mitoma were even more evident than in the healthy cell.' (402)
Figs. 10 and 12-16 in text:
'the threads of the cell network do not stain very deeply with the acid dye, and the bright refraction of the granules blinds the eye to the presence of the threads. It is for this reason that Heidenhain's iron-haematoxylin used with thin sections is so valuable a process. The granules are stained opaquely in shades of black and grey so that their refraction is to a great extent destroyed, and the mitoma itself takes up the iron. This it does to an extent varying not only in different cells, but in different parts of the same cell. With this method one finds then that the eosinophile granules are not isolated bodies lying in a structureless matrix, but that they are greatly enlarged microsomes of an orderly mitoma. I have found this to hold good of all the eosinophile cells which I have seen, both in the blood and elsewhere, and in all the animals I have examined (Figs. 10, 12, 13, 14, 15, 16). The size of the granules is very far from being the same even within the limits of the same cell, although some authors have maintained that in each cell the granules are of the same size. The mistake has arisen from looking only at the entire cells, and the peripheral granules are often very nearly the same in size. A glance however at my Figs. 10, 12, 16, will show how great the range of difference is. Heidenhain believes that the granules are arranged radially to the astrosphere, with the smallest granules next the sphere, the largest at the periphery; and he has pointed out, which I can fully confirm, that there are never any granules within the astrosphere itself The microcentre does not differ from that of other leucocytes, but the limits of the astrosphere are always very evident because of the granules which surround it (Figs. 10, 12,16). The radii within the sphere are usually pretty well stained by the iron-haematoxylin. I have tried to make out Heidenhain's radial arrangement of the granules but have not satisfied myself that it really exists. Appearances are often seen (as in Fig. 16) which suggest it, but I believe that here as in the other leucocytes of which I have spoken the mitoma forms a true network or sponge. There certainly are constantly connecting threads running between the microsomes; I have taken the greatest pains in the drawings only to insert those threads of which I could be certain.' (400-401)
-
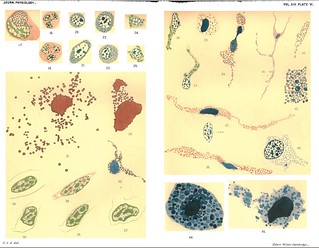 CitesPlate VI, Journal of Physiology 19 (5-6) (1896). Figs. 17-45 from G.L. Gulland, 'On the Granular Leucocytes'.
CitesPlate VI, Journal of Physiology 19 (5-6) (1896). Figs. 17-45 from G.L. Gulland, 'On the Granular Leucocytes'.
Description:Explanation of Plate VI (figs. 17-45):
Fig. 17. Hyaline leucocyte from newt's blood which has ingested the detached pseudopod of an eosinophile cell. Ehrlich-Biondi.
Fig. 18. Oxyphile cell from human blood, with polymorphous nucleus, microcentres in middle of cell. Ehrlich-Biondi.
Fig. 19. As 18, rather larger cell.
Figs. 20-22. Small basophile leucocytes from human blood (leucocytheemia). Methylene-blue.
Fig. 23. The same, but showing the microcentre.
Figs. 24-27. Small basophile cells from the mesentery of an adult rabbit. Methylene-blue.
Fig. 28. Round basophile cell from mesentery of guinea-pig. Nucleus deeply stained. Methylene-blue.
Fig. 29. Amoeboid basophile cell from mesentery of guinea-pig. The cell is passing through a narrow opening a = anterior end. Metbylene-blue.
Fig. 30. The same. c.t.n. = nucleus of one of the connective tissue cells, across which the basophile cell is passing, to show the difference in character of the nucleus.
Fig. 31. Large flat basophile cell from mesentery of rat. Methyleneblue.
Fig. 32. The same, mitoma more evident.
Fig. 33. Section through a basophile cell in the heart-wall of the newt; nucleus deeply stained. Methylene-blue.
Figs. 34-37. Nuclei of basophile cells from peritoneum of newt; granules omitted. Ehrlich-Biondi.
Fig. 38. Nucleus of connective tissue cell from peritoneum of newt. Ehrlich-Biondi.
Fig. 39. Section of basophile cell from heart-wall of newt. Nucleus deeply stained. Methylene-blue.
Fig. 40. Nucleus and part of cell-body of a young basophile cell from peritoneum of newt. Methylene-blue.
Fig. 41. Basophile cell from peritoneum of newt. Methylene-blue, x 200.
Fig. 42. Mitoma from part of Fig. 41, marked 42.
Fig. 43. Mitoma from part of Fig. 41, marked 43.
Fig. 44. Degenerated basophile cell from mesentery of newt. Methyleneblue.
Fig. 45. The same; a later stage.' (416-417)
Fig. 17 in text:
'The mitoma in the eosinophile cell is especially well seen in those of the newt's blood. These cells are markedly amceboid and have the habit of throwing out circular pseudopodia which are often counected to the main part of the cell only by a very delicate thread. Two such cells are shown in Figs. 3 and 6, and it is evident that the threads are often broken through and the spherical portion of the cell-body set free, as the blood contains a large number of them. The mitoma in these is often very clear, as may be seen from Fig. 5 and from Fig. 17, where one of these bodies has been ingested by a "hyaline" leucocyte.' (401)
Figs. 18-19 in text:
''It must not... be thought that the hyaline cell-body is structureless. The constant presence of the microcentre alone would prove the contrary, and though no hint of structure can be seen in the lymphocytes, or even in the smaller hyaline cells, yet as they become larger, one can see the ordinary network of protoplasm (see Figs. 7 and 9). In the hyaline cells the nucleus may remain round or may alter its shape according to Heidenhain's law, that is, according to the relation of its diameter to the diameter -of the whole cell (Figs. 1, 7, 9). The process of change from the small hyaline cell to the finely granular oxyphile cell is associated with an increase in the visibility of the mitoma. It is this, in the early stages, rather than the actual appearance of granules that gives the finely granular appearance to the cell-body. Along with this goes the deformation of the nucleus by amoeboid movements, and the ordinary polymorphous form is soon reached. When these forms are seen in the blood the cells are generally approximately spherical, and the microcentre is situated in the middle of the cell (Figs. 18 and 19). The centrosomes are small, too small for one to be sure of their number as a rule, and I have found them stain better with the unmodified Ehrlich-Biondi stain than with iron-haematoxylin. The limits of the astrosphere are not usually well-marked, except sometimes in the larger cells (Fig. 19).' (399)
'Minute as these microsomes are it can already be made out that they vary in size to a certain extent; on the whole, the largest microsomes are towards the periphery of the cell. In the larger cells of this stage the interstitial substance between the threads of the mitoma becomes increased in amount and the strands are consequently placed further apart. The difference is seen in the contrast between Figs. 18 and 19 and in Figs. 2 and 8.' (399)
Figs. 20-27, 30-38 and 40 in text:
'It is often totally impossible to see the nucleus of the large basophile cells in the mesentery when the specimen is stained with methylene-blue, the stained granules are crowded so closely round the nucleus as to hide it altogether. (Figs. 31 and 32.) One may however in these cases overcome the difficulty by using the Ehrlich-Biondi stain, or by cutting sections of the folded mesentery. I regard the history of the nucleus as being somewhat as follows. In the youngest cells, either in connective tissue (Figs. 24-27) or in blood (Figs. 20-23), the nucleus presents no difference from any ordinary leucocyte-nucleus; the chromatin is arranged in lumps and threads as usual; if anything, the spaces between the threads of the nuclear network are rather larger than usual. As the nucleus becomes older the nuclear reticulum becomes closer and contains relatively more chromatin (Fig. 40); if it passes into amceboid movement or becomes older still (i.e. the cell becomes larger) the staining with methylene-blue becomes more and more diffuse until at last no structure can be made out (Figs. 28, 30, 33). On analysing this-with the Ehrlich-Biondi stain, one finds that this diffuseness of staining is due to the presence of large masses of "basichromatin," staining green, while the amount of pink-stained material, whether linin or "oxychromatin" is very small, and no pink nucleoli are to be seen. I have drawn several such nuclei in Figs. 34-37, and have placed beside them as a contrast the nucleus of a young connective tissue cell (Fig. 38), from the same preparation, which shows well-marked nucleoli and a much more open arrangement of the reticulum. Reinke [note: '(24)' ['Reinke, F. Arch. f. Mikr. Anat. XLIII. 1894.']] has found mitosis once, but only once, in basophile cells. I myself have never been fortunate enough to see it. This extreme rarity of mitosis, taken along with the enormous number of basophile cells in the body, and the relative frequency of degenerated forms, goes far to disprove Kanthack and Hardy's contention that the coarsely granular basophile cells form an isolated group, and to render it almost self-evident that they must be derived from the smaller cells of the same kind, and that these again must be derived from the lymphocytes.' (404-405)
Figs. 20-32, 41-42, 44-45 in text:
'As the cells become larger the granules, as a rule, but not invariably, also grow larger, and become, generally speaking, more uniform in size. I have never seen a cell, however, in which all the granules were quite uniform, though many authors postulate this for all large basophile cells. In all the drawings I have given I have taken the greatest care not only to put the granules in the exact position in which I saw them, but to represent their relative size, and in Figs. 20-32, 41, 42, 44 and 45 (the two latter being degenerated basophiles) I have tried as far as possible to give their colour also. For while it is in these large cells that the most marked metachromatism is to be found, it is to be noted, that in very few cells are the granules stained uniformly red. Some of the very large cells in the rat's mesentery form an exception, but it is more usual to find some granules bright red, others blue, and to find between these all shades of purple. Still the prevailing effect is red.' (407)
Figs. 21-27 in text:
'I shall best be able to describe the changes through which they pass by tracing the cells from the original lymphocyte or hyaline transitionform, from which they are derived. The first change consists not so much in an enlargement of the microsomes as in the superior visibility of the mitoma. As in the eosinophile cells the cell-body becomes larger, and as there the mitoma takes up acid stains, so here it takes up basic stains (Fig. 4). As the cell increases in size, the mitoma retains its visibility and the microsomes become larger and assume the ordinary appearance of granules. These are at first few and scattered irregularly through the cell: some microsomes have increased in size, others have not; the mitoma is always most visible near the microsomes which have taken up the stain. At this stage, illustrated by Figs. 21-23, from the blood, and Figs. 24-27, from the mesentery, the granules always differ markedly in size, much more so than at later stages. They are always uniform in staining, and curiously enough, the very small ones do not often show the metachromatic reaction with methylene-blue. The cells in the blood are generally spherical, but those in connective tissue soon show an irregularity in shape suggestive of amoeboid movements; they put forth those long flat processes which are so characteristic of the larger cells. In Figs. 26 and 27, for instance, all the granules to be seen in the cell are shown, and they were all lying almost in one plane, so that hardly any focussing was required to see them. They must therefore have been lying in a very thin layer of protoplasm, and from the constancy of this appearance we may infer that the spaces through which these leucocytes have to pass between the planes of connective tissue in the mesentery must be very shallow. The rounder cells are generally found in places where, as near the larger blood vessels, the mesentery becomes thicker and allows more space; the elongated cells are generally in the thin parts of the structure.' (406)
Fig. 22, 29-31, 33 and 41-42 in text:
'It has often been remarked that the large cells show a great tendency to leave their granules behind them, thus one might come on a group of granules while the nearest cell was far away. Ballowitz [ref: '(2)' [Ballowitz, E. Anat. Anzeig. vi. 1891.']] was, I think, the first to declare that all or most of these groups of granules were attached to the cell by fine protoplasmic bridges. It is not indeed always easy to show this; in the cell shown in Fig. 31, which was so isolated that there could be no doubt that all the granules represented belonged to it, no trace could be made out of threads extending from granule to granule. They were probably stretched too much to allow them to be visible. Fig. 22, however, taken from the rat also, but not from the same individual, shows the mitoma very clearly, probably because the sheet of cell-substance has not been so greatly stretched. In the elongated forms one often sees narrow bridges, like those shown in Figs. 29, 30, 33, 41, which connect two larger masses of the cell-body; these often consist of a row of granules with a single thread running between (Figs. 29 and 42). As I shall point out when discussing the degeneration of these cells, I have little doubt that when that stage is reached these bridges are torn across, and the granules are actually left behind. This forms an exact parallel to what happens in the eosinophiles of the newt's blood.' (407)
Figs. 23-24 in text:
'Heidenhain has discussed the relation of the centrosomes to basic dyes, and has come to the conclusion that they have no very special affinity. He admits that they do stain with some basic dyes; Van der Stricht, Hermann, and Flemming have stained them with saffranin and gentian-violet, but he himself had negative results both with these and with methyl-violet, methyl-green and thionin. He does not mention methylene-blue. With this they certainly do stain, not in such a way as to constitute a specific reaction, but well enough to allow one, in favourable cases, to make out the relations. Figs. 23 and 24 show the centrosomes, one in a leucocyte from the blood, the other from the mesentery of the rabbit. Neither cell presents any difference of morphological characters from what one might see in an eosinophile cell; change the colours, and no one would know the difference. The distinction in staining between the centrosome and the granules of the cell is slight but distinct; the granules are blue-green, while the centrosomes are rather of an apple-green, of very nearly the same shade of colour as that which the hemoglobin in the red corpuscles assumes when treated with methylene-blue.' (405-406)
Figs. 24-25, 29-32 and 41 in text:
'Westphal [ref: '(4)' ['Ehrlich, P. Farbenanalytische Untersuchungen &c. Gesammelte Mittheilungen. Pt. I. 1891.']] distinguished three main forms of the large basophile cells, flat, spherical, and spindle-shaped, with two or more processes; he remarks however that this is purely a classification for convenience, as transition-forms of every kind are to be seen also. This I can fully confirm, as I can also his dicta that "in individuals of the same species and same age the distribution, number and size of the cells are similar in similar organs," and that in individuals of different species there are great differences in the number &c., of the cells in the same organs. I may add that certain species show a tendency to have cells of one or other shape or size. The mesentery of the rabbit, for instance, contains, speaking generally, small cells, mostly spherical (Figs. 4, 24, 25); of the guinea-pig, larger spindle-shaped cells (Figs. 29 and 30); of the rat, enormous round and flat cells (Figs. 31, 32); while that of the newt has huge spindle cells which send off often several secondary branches from the primary ones (Fig. 41).' (403-404)
Figs. 26-27, 29-30 and 41 in text:
'I consider, contrary to Kanthack and Hardy, that the basophile cells are amoeboid. They examined only the basophile cells of the peritoneal fluid. I find, however, that most of these cells are degenerated. The evidence for their amceboid power is the irregularity of their shape in sections of the tissues (cp. Figs. 26, 27, 29, 30, 41), the phenomenon of "Körnchenzerstreuung," and the way in which the cells are scattered throughout the body. It seems to me that such shapes can only be explained by amoeboid action. In drawing them they must of course be represented in one plane, but one does not in fact find them arranged in that way. The process of Fig. 30 was turned up into quite another direction. Fig. 29 is the most interesting; the end marked a is evidently anterior, and the leucocyte was seen to have been fixed in the act of passing through a narrow hole between two bundles of connective tissue. The nucleus shows in its anterior half that diffuse staining which Arnold long ago pointed out as accompanying amoeboid movement, and which probably depends on the squeezing together of the chromatin-containing filaments. ' (403)
Fig. 33 in text:
'I have spoken as if the elongated basophile cells extended in one plane only, but though this is the case in the membranes, it is not so when these cells occur in ordinary connective tissue. In the heart of the frog or newt, for instance, the branches run in every direction. Fig. 33 will give some slight idea of this.' (408)
Fig. 43 in text:
'In the large basophile cells I have not been able with certainty to make out the position of the microcentre. A frequent appearance is that represented in Fig. 43, where a single large granule forms a centre for a number of smaller ones.' (406)
Figs. 44-45:
'In the portion of cell-body which remains round the nucleus the granules are generally large; the threads of the mitoma lose their power of taking up stains, and the cell assumes the form of a more or less disc-shaped mass of granules, which not only stain badly, but which lose their regular outline. It is curious that in all the markedly degenerated basophile cells which I have seen the granules stained blue with methylene-blue and seemed to have lost the red metachromatism. In Fig. 45 this is well seen. Some of the granules in Fig. 44 retain the red colour to a certain extent, but the drawing rather exaggerates the amount of red in them.' (409)
-
Cited byW.B. Hardy, 'On the Structure of Cell Protoplasm: Part I', Journal of Physiology 24 (2) (1899), pp. 158-210.
Description:'Special mention might be made here of a criticism based upon the structure of oxyphil wandering cells after fixation. On the ground that the granules in these cells form the nodal points of the network Gulland [note: 'This Journal, xix. p. 385. 1896.] claimed that they could not be of the nature of secretory granules, as had been urged by Hankin, Kanthack, and the writer. If these granules really are secretory granules they would according to Gulland occupy the position of paraplastic matter - namely the meshes of the net. The nature of this criticism serves to bring into prominence the shifting artificial nature of the structure in fixed cells, for, as we have seen, the secretory granules of the alveolar cells of the frog's pancreas form the nodal points of the network, while the secretory granules of the orbital gland lie sometimes on the net, sometimes in the meshes, according to the nature of the fixative.' (204)

