- Creation
-
Creator (Definite): Sir Charles Scott Sherrington
- Current Holder(s)
-

- No links match your filters. Clear Filters
-



-
 Cites
Cites Rotating Binocular Lamp
Rotating Binocular Lamp
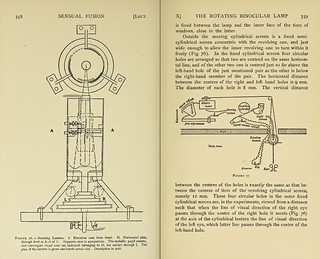
Description:A double sheet of thick milk-glass is observed by transmitted light given by a lamp. This lamp is set in the axis of a rotating cylinder (Fig. 76). In the side of the cylinder are three horizontal rows of rectangular windows, tier above tier. The lamp, though fixed in the axis of rotation of this revolving cylindrical screen, is entirely free from all attachment to it. The milk-glass plate is fixed between the lamp and the inner face of the tiers of windows, close to the latter.
Outside the moving cylindrical screen is a fixed semi- cylindrical screen concentric with the revolving one, and just wide enough to allow the inner revolving one to turn within it freely (Fig. 76). In the fixed cylindrical screen four circular holes are arranged so that two are centred on the same horizontal line, and of the other two one is centred just so far above the left-hand hole of the just mentioned pair as the other is below the right-hand member of the pair. The horizontal distance between the centres of the right and left hand holes is 9 mm. The diameter of each hole is 8 mm. The vertical distance between the centres of the holes is exactly the same as that between the centres of tiers of the revolving cylindrical screen, namely 1 1 mm. These four circular holes in the outer fixed cylindrical screen are, in the experiments, viewed from a distance such that when the line of visual direction of the right eye passes through the centre of the right hole it meets (Fig. 76) at the axis of the cylindrical lantern the line of visual direction of the left eye, which latter line passes through the centre of the left-hand hole.
This being so, the images of the lower left-hand hole and of the upper right-hand hole fuse visually to singleness. They then appear as the middle one of three arranged vertically one above the other.
A black vertical thin screen set at right-angles to the plane of the forehead is introduced (Fig. 77) between the eyes and the holes so as to screen from the left eye all view of the right-hand holes, and from the right eye all view of the left-hand holes.
The revolving screen is driven by an electromotor. The speed of revolution of this motor is controlled by a coarse ad- justment and by a fine adjustment The speed of rotation of the cylindrical screen is recorded by marking the completion of each revolution of its spindle by an electromagnetic signal writing on a travelling blackened surface (Fig. 77). On the same surface the time is recorded by a writing-clock marking fifths of seconds.
The inner revolving screen by its revolution opens and shuts alternately for equal periods the circular holes in the fixed outer screen. The inner screen with its three tiers of windows is made in three pieces, each containing one tier of the windows. The piece containing the middle tier of openings is jointed in such a way that its openings can be set at any desired interval with the openings of the lowest tier. The highest tier is similarly jointed to the middle tier. In this way it can be arranged that the uppermost circular hole is open when the lower ones are closed, or is shut when the lower are closed, or is opened to any desired degree either before or after the lower; further, by removing the top gallery of the rotating screen it can be left permanently open. A similar relationship is also allowed between the middle holes and the lower.
By wearing weak prisms with their base-apex lines vertical the images of the right-hand and left-hand holes can be brought to the same horizontal levels. The observer can then immediately fuse the four images to two by convergence. A horizontal fine thread halving each of the two middle holes, and similar but vertical threads halving the other two holes, serve to certify binocular vision to the observer. When the four holes are all allowed to act thus under appropriate convergent binocular gaze they are seen by the observer as two evenly lighted discs, one vertically above the other, and each cut into quadrants by a delicate black cross. By separately adjustable shutters any one, or any vertically edged fraction of one, of the discs can be separately screened out of vision.' (357-361)
-
 Cited byR.M. Brain, 'Materialising the Medium: Ectoplasm and the Quest for Supra-Normal Biology in Fin-de-Siècle Science and Art', in A. Enns and S. Trower (eds.), Vibratory Modernism (Basingstoke: Palgrave Macmillan, 2013), pp. 115-144.
Cited byR.M. Brain, 'Materialising the Medium: Ectoplasm and the Quest for Supra-Normal Biology in Fin-de-Siècle Science and Art', in A. Enns and S. Trower (eds.), Vibratory Modernism (Basingstoke: Palgrave Macmillan, 2013), pp. 115-144.
Description:'In the 1890s the nerve junction was still understood through the amoeboid theory of neuronal mobility, which held that neurons made functional contacts through pseudopodal movements of the protoplasm of nerve cells. [note: 'Sandra E. Black, 'Pseudopods and Synapses: the Amoeboid Theories of Neuronal Mobility and the Early Formation of the Synapse Concept. 1894-1900', Bulletin of the History of Medicine 55 (1) (1981): 34-58.'] The theory grew in part from the visual evidence: improved microscopy had revealed a new image if neurons as individual cells, with limited movements of extension and contraction that seemed to facilitate inter-neuronal conduction at the 'synapse' (the term itself, from the Greek for 'clasp', was coined by Charles Sherringotn to express this amoeboid movement). [note: 'Charles S. Sherrington coined the term 'synapse' for sections he authored in the seventh edition of Foster's Textbook of Physiology (1997) [sic] as an apt illustration of the amoeboid theory. Interestingly, the term synapse stood, even when the view of the neuronal junction changed, largely through Sherrington's own efforts. To compare Sherrington's amoeboid hypothesis and his later, canonical account of the synapse as a fixed, transverse membrane, see C.S. Sherrington, The Integrative Action of the Nervous System (New York: Scribner's Sons, 1906),']'
-

-
Quoted byJ. Crary, Suspensions of Perception: Attention, Spectacle, and Modern Culture (Cambridge, MA. and London: MIT Press, 1999).
Description:'Sherrington detailed how an organism's functioning involved the ceaseless transformation of perceptual information into purposeful action about within the world. His emphasis on the kineto-temporal character of perceptual experience effectively annuls the notion of seeing "images" of the world, and uses (as did [William] James) the word "stream" to describe how the flow and direction of energy within the nervous system change from instant to instant. At the same time he also employs the (Baudelairean) image of the kaleidescope and other nineteenth-century technological imagery to suggest how changes in the contents of a perceptual field are never just incremental alterations but involve a total reorganization of response:
As a tap to a kaleidescope, so a new stimulus that strikes the receptive surface causes in the central organ a shift of functional pattern at various synapses... The gray matter may be compared with a telephone exchange, where, from moment to moment, though the end points of the system are fixed, the connections between starting points and terminal points are changed to suit passing requirements, as the functional points are shifted at a great railway junction. In order to realize the exchange at work, one must add to its purely spatial plan the temporal datum that within certain limits the connections of the lines shift from minute to minute.
This remarkable hybrid attempt to conjure a suitable four-dimensional image is meant to illustrate the uninterrupted modulation in which certain regions of the nervous system are "shut out" while "vast other regions" are called into play, all in the interest of maintaining the ongoing unity of the individual and of supporting what Sherrington refers to as "the great psychical process of attention." (349-350)
'Sherrington made the fundamental distinction between what he called "distance receptors," which included vision, hearing, and smell, and "immediate receptors," which were the senses of taste and touch. Vision, as part of the evolution of biological survival mechanisms, was the primary "distance receptor" which allowed an organism to extend the limits of its subjective experience beyond its physical boundaries.' (351)
'For Sherrington human vision could not be considered in isolation from the intricate relation of motor behaviour to the "137 million separate 'seeing' elements spread out in the sheet of the retina." The eye moving across a visual field was, he showed, dynamically interconnected "with wide tracts of the musculature as a whole."' (351)
-
Quoted byT. Quick, 'Disciplining Physiological Psychology: Cinematographs as Epistemic Devices, 1897-1922', Science in Context 30 (4), pp. 423-474.
Description:Page numbers refer to 2nd edition (1947).
'This paper considers the significance of what became one of the most emblematic sources of analogy in Bergson's philosophy - that of the cinematograph - for physiological and psychological endeavour during the first decades of the twentieth century. In so doing, it suggests that, by 1922, the scientific landscape in which Bergson had placed so much hope had shifted under his feet. The incorporation of cinematographic devices into physiological and psychological investigation, it suggests, played a critical role in the re-organization of relations between physiology, psychology, and philosophy at this time. Indeed, Bergson's generous comments at Oxford regarding biology and psychology may well even have contributed to this trend. In 1913, Oxford appointed a new Waynflete Professor of Physiology, Charles Scott Sherrington. In 1906, a year before the emergence of Creative Evolution, Sherrington had published a monograph, The Integrative Action of the Nervous System, that would become an exemplary study to twentieth-century physiologists. [note: 'On Sherrington see Smith 2003; idem 2001b; idem 2000; idem 1992, 179-190; Swazey 1969, esp. 1-29.'] Notably, the final chapter of this work centered on a set of experiments that adapted cinematographic devices to very different purposes than those Bergson suggested typified scientific endeavour.'
'In Integrative Action, Sherrington conveyed a holistic conception of nervous action that centred on a physical interpretation of the interaction between nervous elements. Cells featured particularly prominently in Sherrington's analysis. The book's opening words declared that 'nowhere does the cell-theory reveal its presence more frequently in the very framework of the argument than at the present time in the study of nervous reactions.' Furthermore, he suggested, 'the progress of natural knowledge' had enabled biology to pass 'beyond the confines of the study of merely visible form, and... [turn] more and more to the subtler and deeper sciences that are branches of energetics' (Sherrington [1906] 1947, 1).'
'After a consideration of Gaskell and Hering's metabolic propositions, Sherrington devoted nearly three pages of Integrative Action to Macdonald's conclusions. Summarizing the latter's research as leading to the proposition that 'inhibition is the condition in which the possibilities of free motion are most reduced', he suggested that Macdonald's views were 'fertile in suggestion for future experiment' (Sherrington [1906] 1947, 197-200. Quotes on 199-200). Such enthusiasm reflected a new emphasis within physiology on the study of nerve junctions themselves as sites at which nerve conduction might be interrupted. In his Textbook of Physiology of 1900, Edward A. Schäfer had suggested that impulses were 'momentarily arrested at these places of contact of nerve cells with one another', which he referred to as 'synapses' (Schäfer 1900, 608). Sherrington, characterizing such arrests as manifestations of a 'neurone threshold', identified it unequivocally with a non-contiguous, synaptic transmission:
at each synapse a small quantity of energy... acts as a releasing force to a fresh store of energy not along a homogeneous train of conducting material, as in a nerve fibre... but across a barrier which whether lower or higher is always to some extent a barrier (Sherrington [1906] 1947, 157).
This insistence on the functional significance of barriers between cells marks a critical change of emphasis within physiological considerations of inhibitory mechanisms. Where explanations of variations in the rate of electrical transmission along nerves had centred on cells and the substances that composed them, Sherrington and his colleagues characterized such variability as dependent on changes in the thresholds at which transmission between cells could be effected. Neither cell bodies nor protoplasm were the most physiologically significant conditions for variation in the transmission of nervous impulses: rather, 'synapses' were.'
'Despite his enthusiasm regarding McDougall's explanation of 'reciprocal innervation', Sherrington's characterization of it limited its explanatory scope to a particular aspect of a broader, physical, phenomenon. He admitted that McDougall's conclusions presented 'an explanation for the transition from one antagonistic reflex to another' (Sherrington [1906] 1947, 200). As a framework for inhibition, however, they tended
to sever... central nervous inhibition – of which I regard reciprocal innervation of antagonistic muscles as but one widely spread case – from other forms.... It appears to me unlikely that in their essential nature all forms of inhibition can be anything but one and the same process (Sherrington [1906] 1947, 203).
Other forms of inhibition, a uniformly-caused function of nervous action, could not be explained by McDougall's scheme. McDougall's psychological insights were re-cast in Integrative Action as contributions to the experimental study of physical (rather than mechanical) nervous function.'
'In Integrative Action, Sherrington introduced his cinematographic studies as pertaining to the question of whether or not it was possible to extend his conclusions regarding nervous activity in simple reflexes to the study of sensations. For the majority of the book, he had, as he noted, supposed the animal 'a puppet without passions, memory, feelings, sensations, let alone ideas concrete or abstract.' Now, in his final chapter, he queried whether 'we [can] at all compare with the simultaneous co-ordination of the nervous factors in a motor reflex the synthesis of the nervous elements whose combination underlies a simple sense-perception?' (Sherrington [1906] 1947, 353, 355) Though he notably did not go so far as to make any positive claim, he thereby intimated that complex psychological phenomena might be explained by reference to his physical interpretation of nervous action. [note: 'For an account of Sherrington's conception of sensation as it emerged later in his career see Smith 2001a, 232-235.'] Less clear from Sherrington's text, however, is that were such a claim to have been established, it would have turned back a tide of opinion that had been gaining ground in physiology since at least the middle of the nineteenth century.'
'Sherrington's studies sought to bring the production of cinematographic effects of continuity within the purview of his physical physiological schema. His goal was not merely to arrive at an accurate estimation of the rate at which flickering sensations transformed into continuous ones, but also to interrogate the relationship between the retina, the nerves, and visual sensations more generally. Hering's studies had suggested that the optic nerves had to connect before they arrived at the centre of consciousness, as it was in the eyes and their associated nerves that vision was produced. Helmholtz's insistence that knowledge was a judgemental construct, in contrast, seemed to imply that all sensory organs were connected with the cerebrum individually - it was only through the mind bringing individual sense-experiences together than external perception was generated. By creating a situation in which 'corresponding' points on the retina could be stimulated in similar but crucially alterable ways, Sherrington sought to establish with greater surety than had McDougall the modes of interaction between visual fields. By blocking off some of the holes and one or another of the rectangles cut into the rotating screen, it was possible to create situations in which retinal points could be stimulated either simultaneously or alternately, but always at the same rate. Thus, Sherrington hoped, it would be possible to examine whether stimulatory effects were 'additive' between corresponding nerves, or whether McDougall's contentions regarding their 'mutual inhibition' were in fact accurate (Sherrington [1906] 1947, 375-378). If binocular visual sensation could be shown to be additive, Helmholtz's conclusions could be confirmed. If they inhibited one another, Hering would be supported. Here then appeared a crucial test of a long-standing dispute within physiological psychology.
Despite the lengths to which Sherrington went to ensure that both his subjects and his objects were as carefully arranged and closely controlled as possible, however, he was unable to reach any firm conclusions regarding his results. Sherrington's aim had been to evaluate the extent to which his conclusions regarding the physical nature of simple reflex action could be brought to bear on the interpretation of complex sensory phenomena. But when it came to drawing conclusions from these studies, he himself wavered between the two above-noted possibilities. The problem was not - as had hampered so many prior studies in this vein - disagreement amongst individual observers. Rather, it concerned the seemingly contradictory nature of the results themselves. Whether retinal stimulations could be said to sum together or mutually interfere depended, it seemed, on the mode of stimulation: ''corresponding retino-cerebral' points', he perplexedly concluded, 'retain individuality as regards time-relations... [but are] completely confluent by reference to visual space' (Sherrington [1906] 1947, 366). Furthermore the 'rule of combination' revealed by the experiment 'finds little solution by appeal to summation or interference of retinal and purely physiological processes' (Sherrington [1906] 1947, 369). Though Sherrington leant towards a Helmholtzian conception of visual sensation, introspective sensory investigation, when conducted under carefully controlled laboratory conditions, did not lead to clear-cut physiological conclusions.
The possibility that the study of visual illusions could inform study of the extent to which optic nerves actively contributed to sensation had taken Sherrington a long way from his starting point of the physical interaction of individual nerves. Integrative Action continued to portray the study of 'sensual reaction' as at least potentially part and parcel of that of nerve reactions more generally. Although the difficulty of interpreting the flicker experiment showed that 'hasty conclusions' regarding the parallels between simple and 'sensual' reflexes were unwise, it was nevertheless possible to insist on a 'likeness of nervous reactions expressed by muscular and other effector-organs to reactions whose evidence is sensual' (Sherrington [1906] 1947, 384). More generally, the correspondence of sensations to positions in space, to which the body could direct its actions, indicated that 'physiology and psychology, instead of prosecuting their studies... more strictly apart... will find it serviceable for each to give the results achieved by the other even closer heed than has been customary hitherto' (Sherrington [1906] 1947, 385). Nevertheless, Sherrington's cinematographic studies were not a success in his own (physiological) terms, and he would not produce any further work in this vein. By the end of the First World War, the rooms at Oxford that McDougall used for his psychological experimentation had been requisitioned for more strictly physical investigations (Oldfield 1950, 382). Such differentiation of physiological from psychological endeavour would set a trend for these sciences during the first half of the twentieth century.'
-
-

-
Related toMaterial relating to Sir Charles Sherrington's interest in fatigue
Description:from 1961 paperback of 2nd ed. (1947).
On declining nervous response as an aspect of fatigue:
'A spinal reflex under continuous excitation or frequent repetition becames weaker, and may cease altogether. This decline is progressive, and takes place earlier in some kinds of reflexes than it does in others. …
A reflex as it tires shows other changes besides decline in amplitude of contraction...; it becomes tremulous, and the tremor becomes progressively more marked and more irregular... Finally, an irregular phasic tremor of all the muscles is all that remains. It is not the flexor muscles themselves which tire out, for these, when they contract no longer during the flexion-reflex, still contract in response to the scratch-reflex which also employs them.' (215)'… the phenomena are not referable to [(215), (217)] the muscle, for when excited through other reflex channels, ot through its motor nerve directly, the muscle shows its contraction well.' (215-217)
On the role of fatigue in the expression of relfex arc competing for the same 'final common path' (ie. set of motor neurons):
'In the hind limb of the spinal dog the extensor-thrust is in-elicitable during the flexion-reflex... But when the flexion-reflex is kept up by appropriate stimulation of a single point over a prolonged time , so that it shows fatigue, the 'extensor thrust' becomes again elicitable... In other words, it can dispossess the rival reflex from a common path when that rival is fatigued, though it cannot do so when the rival action is fresh and powerful.' (223)
The function of fatigue:
'It seems a process elaborated and preserved in the selective evolution of the neural machinery. One obvious use attaching to it is the prevention of the too prolonged continuous use of a 'common path' by any one receptor. It precludes one receptor from occupying for long periods an effector organ to the exclusion of all other receptors. It prevents long continuous possession of a common path by any one reflex of considerable intensity. It favours the receptors taking turn about. It helps to insure serial variety of reaction. The organism, to be successful in a million-sided environment, must in its reactions be many-sided. Were it not for [(224) (225)] such so-called 'fatigue', an organism might, in regard to its receptivity, develop an eye, or an ear, or a mouth, or a hand or leg, but it would hardly develop the marvellous congeries of all those various sense-organs which it is actually found to possess.' (224-225)
On the relevance of fatigue to the psychology of the senses:
'If instead of motor effects sensual are examined, analogous phenomena are observed. A visual image is more readily inhibited by a competing image in the same visual field when it has acted for some time than when it is first perceived (W. McDougall).' (224) [n. to McDougall, W. (1901) Mind, x, no. 37.]
-
Related toThe British Association, Opening Address by Prof. C. S. Sherrington, Nature, 8 Sept 1904 (70), pp. 460-466.
Description:The speech outlines the primary points made in the book, often employing exactly the same wording as the latter.


