Tag: logwood
No results
|
|
|
cites |
 |
|
  |
|||
|
Explanation of Plate XXXI (figs. 1-5): 'Fig. 1. Contents of experimental chamber that had remained 72 hours in the peritoneal cavity of the rabbit. Five large amoeboid plasma-cells, with altered red corpuscles and apparently dead leucocytes. Outlined with camera lucida. Apochromatic oil immersion and ocular No. 4. Zeiss. Prepared over osmic vapour. Fig. 2. Contents of a chamber for 18 hours in the peritoneal cavity (rabbit); near the centre of the chamber. Fibrin filaments, leucocytes, red corpuscles, and an ill-defined granular mass forming a nodal point in the fibrinous network-the beginning of a "cell-islet." Outlined under camera. Similar method of preparation, and similar magnification to preceding. Fig. 3. Fragment of inflammatory membrane formed within a chamber placed for three days in the subcutaneous tissue (guinea-pig). Islets and groups of islets scattered through the membrane. Zeiss, Obj. A, Oc. 2. Osmic acid solution, and Ehrlich's logwood. Fig. 4. Contents of same chamber as in Fig 1. Close to the opening of the chamber. Five plasma-cells, one of them continuing a leucocyte within a large vacuole. Magnification and method of preparation as in Fig. 1. Fig. 5. Contents of same chamber. Two plasma-cells and two red corptiscles; the plasma-cells are indistinguishably united with fine filaments of fibrin in their surrounding, some of which are given in the figure. Osmic acid vapour. Zeiss, apochr. system, oc. No. 2.' (575)
Figs. 1 and 4-5 in text: 'The preparations gave an almxiost bewildering number of examples of the infinite variation in shape of the large amoeboid plasma-cells, which also varied very considerably in size, and as to granules. The body of the cell was for the most part plate-like, being in many instances extended into so thin a film that its exact limit was hard to determine, especially when, as occasionally happened, the granules of the cell-body were less pronounced towards the periphery. Some idea of the wide diversity of outline exhibited by individual cells may be gathered from our figures. Cf Figs. 1, 4, 5, 6, 7 and 8, Plates XXXI. and XXXII.' (558) 'In the specimens obtained from chambers that had rested for seventy-two hours in the subcutaneous tissue of the guinea-pig, we found individuals among the plasma-cells, which showed wellmarked vacuolation, Figs. 1, 4, 5, Pl. XXXI. For the most part the matter within the vacuole was a granular debris that furnished no sufficient clue as to its nature. But in a few it was indisputable that the vacuole contained, more or less altered but still perfectly easily recognisable, a leucocyte or red blood corpuscle. In Fig. 4 is shown the appearance presented by one of these cells. A large vacuole contains a somewhat faintly stained body, which is finely granular and indistinctly nucleated. It is a little smaller than is the nucleus of the plasma-cell itself. Fine threads seemed to pass from the sides of the vacuole across the cavity to the substance of the included leucocyte. Taken with the context afforded by examination of other cells in the neighbourhood we believe that this and other similar instances were examples of leucocytes lying in vacuoles in the plasma-cells. Many stages of ingestion could be found. Cf. Figs. 1, 4, 5, Pl. XXXI. Simple approximation, the hollowing out of a little bay in the side of the plasma-cell into which the leucocyte was as it were drawn, partial inclusion, total inclusion - all these were exemplified.' (559)
Figs. 1 and 5 in text: 'Contiguous plasma-cells or even those a little distance apart were often connected together by their processes (Figs. 1, 5, 7 and 8, Plates XXXI. and XXXII.). The bands of connection might be short thick arms or long gossamer threads of protoplasm. By similar arms and threads the cells seemed to adhere to the most diverse objects in their surrounding. The surface of the cover-glass, a filamllent of fibrin, a hair, a fibre of cotton, a lump of the cement fastening the sides of the chamber together, all afforded points to which the processes from the plasma-cells would cling (Figs. 14 and 15).' (560)
Fig. 1 in text: 'Here must be mentioned another sign of degeneration in the leucocytes examined in these chambers [note: 'Kuss, Paris, 1846. Paget, Surgical Pathol. p. 151.']. Many of them showed the triple and multiple nuclear bodies that are universally regarded as evidence of the lethal disintegration of the nucleus - as Fleming names it, the "fragmentation" of the nucleus. On the other hand the cell-body of the leucocyte was not granular or fatty, but fairly evenly though deeply tinted by the osmium. These points are seen in Fig. 1, Plate XXXI.' (556-557)
Figs. 2-3 in text: 'Eighteen Hours.- In chambers removed after the appearance of fibrin within them, but before the stay within the body had exceeded eight and forty hours, it was usual to find a number of areas in which leucocytes were present in much greater numbers than elsewhere. Fig. 2, Pl. XXXI. The tendency to collect to certain points which the leucocytes evinced in even very early specimens was more marked in these later preparations. About the nodal points of the fibrinous network crowds of them were present. The ouitlying individuals were frequently arranged in lines along the converging filaments of fibrin. The older within certain limits these films of coagulum the more obvious the aggregation of the leucocytes into certain groups. For convenience on account of their prominence. and apparent importance in subsequent stages we have been accustomed to refer to these groups shortly as the cell-islets. Cf. Fig. 3, Pl. XXXI. They are little collections of cells, occurring constantly, scattered about in the thin cellular membranes which grow over and within the glass chambers.' (557)
|
||||||||
|
|
|
cites |
 |
|
|
|||
|
Explanation of Plate XXXII (figs. 6-13): 'Fig. 6. Giant cells from chamber 72 hours in the peritoneal cavity (rabbit). Zeiss apochr. system, ocul. 5. Osmic acid vapour. Fig. 7. Plasma-cells from same preparation which furnished Fig. 6. Fig. 8. " Cell islet " from inflammatory film obtained in a chamber left eight days in the subcutaneous tissue of the guinea-pig, At the margin it is united to outlying plasma-cells. Zeiss oil, oc. 4. Osmic acid vapour. Fig. 9. Young cicatricial tissue of anastomosing branched cells, some of which are represented under the higher magnification in Fig. 17. From a thrombosed artery (syphilis) near the centre of the thrombus. Zeiss A, oc. 3. Logwood. Preparation kindly shown us by Dr Seymour Sharkey. Fig. 10. "Cell islet" from inflammatory membrane obtained from chamber five days in the peritoneal cavity of the rabbit. Osmic acid solution. Zeiss oil imm. and oc. 2. Fig. 11. Mass of blood-cells (? clot) surrounded by fibroblastic cells, and invaded by them at four places. Inflammatory membrane from chamber eight days in subcutaneous tissue. Magnification as in preceding, and prepared in similar manner. Fig. 12. Fusiform plasma-cell (fibroblast) surrounded by a fibrillated material which forms a thread-like band of connective tissue. Zeiss oil and oc. 4. Osmic vapour. From chamber 10 days in subcutaneous tissue. Fig. 13. Similar but larger and thicker fibrous band from same preparation. Similar preparation and magnification.' (575-576)
Figs. 6-8 in text: 'The preparations gave an almxiost bewildering number of examples of the infinite variation in shape of the large amoeboid plasma-cells, which also varied very considerably in size, and as to granules. The body of the cell was for the most part plate-like, being in many instances extended into so thin a film that its exact limit was hard to determine, especially when, as occasionally happened, the granules of the cell-body were less pronounced towards the periphery. Some idea of the wide diversity of outline exhibited by individual cells may be gathered from our figures. Cf Figs. 1, 4, 5, 6, 7 and 8, Plates XXXI. and XXXII.' (558)
Fig. 6 in text: 'There were present also in chambers of eighteen hours', twentytwo hours', twenty-six hours', forty-eight hours', and seventy-two hours' standing, as also in others of older date containing well formed granulation tissue, many giant cells (Fig. 6) - huge multi-nucleate cells, that obviously in many instances were cell-fusions. ' (560)
Figs. 7-8 in text: 'Contiguous plasma-cells or even those a little distance apart were often connected together by their processes (Figs. 1, 5, 7 and 8, Plates XXXI. and XXXII.). The bands of connection might be short thick arms or long gossamer threads of protoplasm. By similar arms and threads the cells seemed to adhere to the most diverse objects in their surrounding. The surface of the cover-glass, a filamllent of fibrin, a hair, a fibre of cotton, a lump of the cement fastening the sides of the chamber together, all afforded points to which the processes from the plasma-cells would cling (Figs. 14 and 15).' (560)
Figs. 8-9 in text: 'In membranes of ten, fourteen, and even eighteen days' growth, not all the cells nor even the majority were spindle-shaped. A vast number were triradiate, and multiradiate; some had but one process; very few were rounded. Many recalled to mind the branched fixed corpuscles of the cornea. Long tapering branches united cell to cell, not only the cells of one plane one with another, but the cells of different planes also (Figs. 8, 9 and 17). A meshwork of infinite variety and complexity was thus established. But in all these examples of plasma cells in the stable as well as in the previously described labile forms, the granular nature of the cell substance and the clear oval nucleus were characters never lost.' (563)
Figs. 8 and 10 in text: 'in the specimens of more than forty-eight hours' duration, the plasma-cells begin to apply themselves to the islet-groups of leucocytes. Cf. Figs. 8 and 10. They surround the leucocytes. The islets come to consist of a central portion made up of leucocytes, and an outer zone of large and granular plasma-cells. In this way the islets seem to increase rapidly in size. Neighbouring islets appear to become merged together.' (561)
Fig. 11 in text: 'It was among the plasma-cells of the fringe of the islets that we noticed the earliest regularly fusiform cells, the immediate precursors of fibrous elements in the new tissue. It is true that plasma-cells of an irregular spindle-shape were observable not rarely among even the earliest of the plasma-cell swarm entering the chamber. But in those instances the outline was probably but one of many which the amoeboid cell successively assumed, and generally it was not of the same character as the regularly fusiform type prevailing among these plasma-cells in the outskirts of an islet. In that latter the majority of the cells lay in lines concentrically set about a core of ill-stained, broken-down matter that composed the centre of the mass. Cf. Fig. 11, Pl. XXXII. The fusiform fibroblasts began in fact the encapsulation of the débris of the breaking-down blood cells, &c.' (526)
Figs. 12-13 in text: 'Older specimens revealed further progress in the formation of a fibrous-tissue membrane. After a stay of eight days, or ten days, or fourteen days in the subcutaneous tissue in many instances the islets consisted of plasma-cells alone. The leucocytes had disappeared. The pigmented remnants of the red blood corpuscles were much longer traceable. In many places along certain lines the spindle-shaped cells had become attenuated, and formed distinct bands and often long and delicate cords (Figs. 12, 13). In many places in the tenth day specimens, and in some of the eighth day ones an inter-cellular substance showing fibrillation exists (Fig. 12).' (562) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
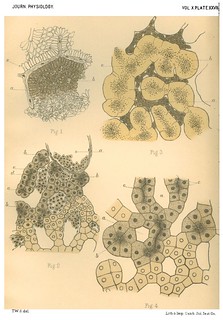
Explanation of Plate XXVII (figs. 1-4): 'Fig. 1. Portion of a section of the liver of the Lamprey. (x about 400 diam.) a. Blood capillary with nucleated endothelial wall. b. Blood capillary cut transversely. Around it is seen a radial arrangement of the elongated liver cells. c. Solid hepatic cylinder with loosely arranged cells. Fig. 2. Part of a section of the liver of the Eel. ( x 400 diam.) a. Transverse section of a larger vessel (probably radicle of hepatic vein). b. Liver tubules cut transversely. c. Lumen of a liver tubule. d. Liver tubule in longitudinal section. e. Nucleated endothelial wall of a capillary blood space. Fig. 3. Section of an osmic acid preparation of the liver of the Frog. (x 400 diam.) a. Blood corpuscles lying in a capillary vessel. b. Outer lightly stained zone of the cells of the liver tubules. c. Inner black-stained granular zoite around the lumen of the tubules. Fig. 4. Section of a logwood stained preparation of the liver of the Frog. (x about 400 diam.) a. Sections of the network of spaces for blood capillaries. b. Liver tubules in transverse sectioln. c. Lumina of liver tubules cut longitudinally.' (426-427)
Fig. 1 in text: 'Our first impression was, that there were in this liver large lumina with cells grouped round them though not in a single layer; but further examination made it clear that these supposed lumina are really blood channels and that between these blood channels the liver cells are disposed in solid anastomosing cylinders, the central cells of which are loosely arranged, with intercellular spaces occupying what may be viewed as the lumen of a potential tube. (Plate XXVII. Fig. 1.) The relative proportion of cell-cylinder to blood-vessel is extremely large, and between any two blood-vessels the number of layers of cells varies from two to five or six. The cells of which the intervascular cylinders are composed naturally fall into two classes, distinguished by their form and arrangement;- (a) those immediately adjacent to the blood-vessels, which have the form and arrangement of a columnar epithelium disposed radially round the blood channels, and each measuring 5µ in width and about 15µ in length, (b) those situated within the cell cylinders, which are generally polyhedral in form, some few being slightly elongated and each measuring about 5µ in all diameters. The latter are disposed in a sponay manner and in hardened specimens show channels running between them.' (414)
Figs. 1-2 in text: 'There is nothing in the knownw history of the development of the liver which opposes the idea that the gland may have arisen from a solid mass of hypoblast cells formed at the extremity of the primitive hepatic diverticulum and which growing and forcing its way into the surrounding mesoblast, becomes itself interpenetrated by ingrowing mesoblast cells. By the development of these latter into blood capillaries the hypoblastic cell mass would become broken up into a network of solid anastomosing rods, in which secretion channels would subsequiently form, and which would be more or less fine according to the completeness with which the ingrowth of developing blood-vessels took place... An examination of the liver of the types we have described in the light of our hypothesis will show that all the appearances can be explained by it. We can find no other satisfactory interpretation of the arrangement seen in the liver of the lamprey (Plate XXVII. Fig. 1). A comparison of a section of the eel's liver with that of a mammal (Plate XXVII. Fig. 2 and Plate XXIX, Fig. 12) shows clearly that a more intimate subdivision by capillaries in the case of the latter would account for the differences, and at the same time it is seen that the relative magnitude of the blood capillaries in the two cases is very different - those of the eel can scarcely be called "capillaries". That this is the true difference in the two cases is clear from a comparison of a section of the liver of an embryo mammal with that of the adult and with that of the eel (Plate XXVII. Fig. 2, and Plate XXIX., Figs. 9 and 12). On examining a section of the liver of the newt, the appearances seen, making allowance for the different sizes of the cells and smaller details, are more like those of the mammal's liver than that of any other of our types - (Plate XXVIII. Fig. 6) - There has been in this case a penetration of the mass of liver cells sufficiently intimate to have left but three or four rows of cells to form the tubules, and at the same time the total vascularity of the organ, as shown by the size of the blood spaces, is not so great as in the case of the eel or the mammal.' (423-424)
Fig. 2 in text: 'The cells of the eel's liver are of medium size, and are granular throughout with a condensation of granules towards the lumen, which is distinct though small. The number of rows of cells forming the wall of the tubule is generally five or six, but in some cases is four. The diameter of the tubule averages 25µ, but is not quite uniform, (Plate XXVII. Fig. 2).' (412)
Figs. 3-4 in text: 'The cells of the frog's liver are large and clear. They have granules on their inner borders, next the lumen, which feature is brought out very clearly by treatment with osmic acid, but is also visible in good logwood-stained specimens, one of which is figured in Plate XXVII. Fig. 4. Langley [note: 'Langley, loc. cit.'] states that in summer a hungry frog has granules equally scattered throughout the cells of its liver, and that during winter there is a marked inner granular zone. We can confirm this, and the specimen figured in Plate XXVII. Fig. 3, is from a winter frog, which on account of the distribution of the granules is the one best suited for tracing a lumen. In an osmic acid preparation the anastomosing liver tubules are rendered very plain by the darker staining of the blood in the capillaries, and they have the appearance of a lightly stained mass with the division into cells not very obvious, but with a well marked inner zone of black granules. In a logwood or cochineal stained specimen the number of rows of cells is seen to be usually five.' (415) |
||||||||
