Tag: cochineal
No results
|
|
|
cites |
 |
|
  |
|||
|
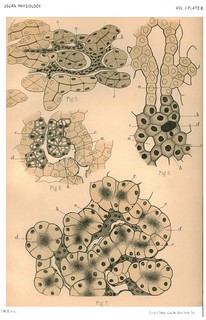
Explanation of Plate XXIX (figs. 9-12): 'Fig. 9. Section of the liver of an Embryo Cat, 15 mm. in length. (x 400 diam.) a. Capillary blood space. b. Endothelial wall of capillary blood-vessel. c. A liver tubule. d. Lunmen of a liver tubule cut transversely. Fig. 10. Portion of an osmic acid preparation of the liver of the Rat. (x about 400 diam.) a. Spaces occupied by the network of blood capillaries cut longitudinally. b. Similar spaces cut transversely. c. Transverse section of a bile capillary lying between two liver cells. d. A similar bile capillary lying at the angle where three liver cells meet. Fig. 11. A similar preparation of the liver of the Mole. (x about 400 diam.) a. Spaces for blood capillaries. b. Section of a bile capillary lying between four cells. A sinmilar one is seen near it, lying between three cells. Fig. 12. Portion of a section of a cochineal stained specimen of the liver of the Pig, from the peripheral part of a lobule. (x about 400 diam.) a. Nucleated wall of a capillary blood-vessel. b. Liver cells showing a collection of granules where two or three cells are contiguous.' (427-428)
Figs. 9 and 12 in text: 'There is nothing in the known history of the development of the liver which opposes the idea that the gland may have arisen from a solid mass of hypoblast cells formed at the extremity of the primitive hepatic diverticulum and which growing and forcing its way into the surrounding mesoblast, becomes itself interpenetrated by ingrowing mesoblast cells. By the development of these latter into blood capillaries the hypoblastic cell mass would become broken up into a network of solid anastomosing rods, in which secretion channels would subsequiently form, and which would be more or less fine according to the completeness with which the ingrowth of developing blood-vessels took place... An examination of the liver of the types we have described in the light of our hypothesis will show that all the appearances can be explained by it. We can find no other satisfactory interpretation of the arrangement seen in the liver of the lamprey (Plate XXVII. Fig. 1). A comparison of a section of the eel's liver with that of a mammal (Plate XXVII. Fig. 2 and Plate XXIX, Fig. 12) shows clearly that a more intimate subdivision by capillaries in the case of the latter would account for the differences, and at the same time it is seen that the relative magnitude of the blood capillaries in the two cases is very different - those of the eel can scarcely be called "capillaries". That this is the true difference in the two cases is clear from a comparison of a section of the liver of an embryo mammal with that of the adult and with that of the eel (Plate XXVII. Fig. 2, and Plate XXIX., Figs. 9 and 12). On examining a section of the liver of the newt, the appearances seen, making allowance for the different sizes of the cells and smaller details, are more like those of the mammal's liver than that of any other of our types - (Plate XXVIII. Fig. 6) - There has been in this case a penetration of the mass of liver cells sufficiently intimate to have left but three or four rows of cells to form the tubules, and at the same time the total vascularity of the organ, as shown by the size of the blood spaces, is not so great as in the case of the eel or the mammal.' (423-424) |
||||||||
|
|
|
cites |
 |
|
|
|||
|
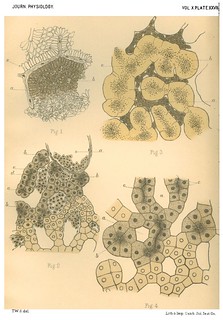
Explanation of Plate XXVII (figs. 1-4): 'Fig. 1. Portion of a section of the liver of the Lamprey. (x about 400 diam.) a. Blood capillary with nucleated endothelial wall. b. Blood capillary cut transversely. Around it is seen a radial arrangement of the elongated liver cells. c. Solid hepatic cylinder with loosely arranged cells. Fig. 2. Part of a section of the liver of the Eel. ( x 400 diam.) a. Transverse section of a larger vessel (probably radicle of hepatic vein). b. Liver tubules cut transversely. c. Lumen of a liver tubule. d. Liver tubule in longitudinal section. e. Nucleated endothelial wall of a capillary blood space. Fig. 3. Section of an osmic acid preparation of the liver of the Frog. (x 400 diam.) a. Blood corpuscles lying in a capillary vessel. b. Outer lightly stained zone of the cells of the liver tubules. c. Inner black-stained granular zoite around the lumen of the tubules. Fig. 4. Section of a logwood stained preparation of the liver of the Frog. (x about 400 diam.) a. Sections of the network of spaces for blood capillaries. b. Liver tubules in transverse sectioln. c. Lumina of liver tubules cut longitudinally.' (426-427)
Fig. 1 in text: 'Our first impression was, that there were in this liver large lumina with cells grouped round them though not in a single layer; but further examination made it clear that these supposed lumina are really blood channels and that between these blood channels the liver cells are disposed in solid anastomosing cylinders, the central cells of which are loosely arranged, with intercellular spaces occupying what may be viewed as the lumen of a potential tube. (Plate XXVII. Fig. 1.) The relative proportion of cell-cylinder to blood-vessel is extremely large, and between any two blood-vessels the number of layers of cells varies from two to five or six. The cells of which the intervascular cylinders are composed naturally fall into two classes, distinguished by their form and arrangement;- (a) those immediately adjacent to the blood-vessels, which have the form and arrangement of a columnar epithelium disposed radially round the blood channels, and each measuring 5µ in width and about 15µ in length, (b) those situated within the cell cylinders, which are generally polyhedral in form, some few being slightly elongated and each measuring about 5µ in all diameters. The latter are disposed in a sponay manner and in hardened specimens show channels running between them.' (414)
Figs. 1-2 in text: 'There is nothing in the knownw history of the development of the liver which opposes the idea that the gland may have arisen from a solid mass of hypoblast cells formed at the extremity of the primitive hepatic diverticulum and which growing and forcing its way into the surrounding mesoblast, becomes itself interpenetrated by ingrowing mesoblast cells. By the development of these latter into blood capillaries the hypoblastic cell mass would become broken up into a network of solid anastomosing rods, in which secretion channels would subsequiently form, and which would be more or less fine according to the completeness with which the ingrowth of developing blood-vessels took place... An examination of the liver of the types we have described in the light of our hypothesis will show that all the appearances can be explained by it. We can find no other satisfactory interpretation of the arrangement seen in the liver of the lamprey (Plate XXVII. Fig. 1). A comparison of a section of the eel's liver with that of a mammal (Plate XXVII. Fig. 2 and Plate XXIX, Fig. 12) shows clearly that a more intimate subdivision by capillaries in the case of the latter would account for the differences, and at the same time it is seen that the relative magnitude of the blood capillaries in the two cases is very different - those of the eel can scarcely be called "capillaries". That this is the true difference in the two cases is clear from a comparison of a section of the liver of an embryo mammal with that of the adult and with that of the eel (Plate XXVII. Fig. 2, and Plate XXIX., Figs. 9 and 12). On examining a section of the liver of the newt, the appearances seen, making allowance for the different sizes of the cells and smaller details, are more like those of the mammal's liver than that of any other of our types - (Plate XXVIII. Fig. 6) - There has been in this case a penetration of the mass of liver cells sufficiently intimate to have left but three or four rows of cells to form the tubules, and at the same time the total vascularity of the organ, as shown by the size of the blood spaces, is not so great as in the case of the eel or the mammal.' (423-424)
Fig. 2 in text: 'The cells of the eel's liver are of medium size, and are granular throughout with a condensation of granules towards the lumen, which is distinct though small. The number of rows of cells forming the wall of the tubule is generally five or six, but in some cases is four. The diameter of the tubule averages 25µ, but is not quite uniform, (Plate XXVII. Fig. 2).' (412)
Figs. 3-4 in text: 'The cells of the frog's liver are large and clear. They have granules on their inner borders, next the lumen, which feature is brought out very clearly by treatment with osmic acid, but is also visible in good logwood-stained specimens, one of which is figured in Plate XXVII. Fig. 4. Langley [note: 'Langley, loc. cit.'] states that in summer a hungry frog has granules equally scattered throughout the cells of its liver, and that during winter there is a marked inner granular zone. We can confirm this, and the specimen figured in Plate XXVII. Fig. 3, is from a winter frog, which on account of the distribution of the granules is the one best suited for tracing a lumen. In an osmic acid preparation the anastomosing liver tubules are rendered very plain by the darker staining of the blood in the capillaries, and they have the appearance of a lightly stained mass with the division into cells not very obvious, but with a well marked inner zone of black granules. In a logwood or cochineal stained specimen the number of rows of cells is seen to be usually five.' (415) |
||||||||
